Kasvikudokset: johtavat, mekaaniset ja erittävät. Johtavia kankaita. Toiminnot ja rakenteelliset ominaisuudet Missä sähköä johtava kudos sijaitsee laitoksessa
Evoluutioprosessissa julkaisun kanssa korkeampia kasveja Maalla he kehittivät kudoksia, jotka saavuttivat suurimman erikoistumisensa kukkiviin kasveihin. Tässä artikkelissa tarkastellaan tarkemmin, mitä kasvikudokset ovat, minkä tyyppisiä niitä on, mitä toimintoja ne suorittavat, sekä kasvikudosten rakenteellisia ominaisuuksia.
Kangas ovat soluryhmiä, jotka ovat rakenteeltaan samanlaisia ja suorittavat samat toiminnot.
Tärkeimmät kasvikudokset on esitetty alla olevassa kuvassa:
Kasvikudosten tyypit, toiminnot ja rakenne.


Kasvien sisäkudos.
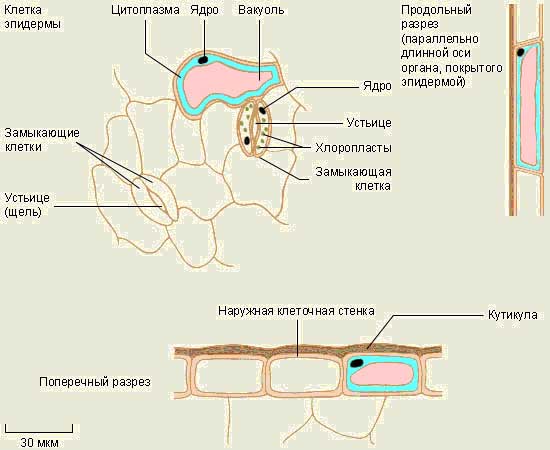


peittää kudosta kasvit - kuori
Johtava kasvikudos.
| Kankaan nimi | Rakenne | Sijainti | Toiminnot |
| 1. Puuastiat - ksyleemi | Ontot putket, joissa on lignified seinät ja kuollut sisältö | Puu (ksylemi), joka kulkee juurta, vartta, lehtisuonia pitkin | Veden ja mineraalien johtaminen maaperästä juurille, varrelle, lehdille, kukille |
|
2. Seula putket niini - floem Mukana olevat solut tai kumppanisolut |
Pystysuora elävien solujen rivi, jossa on seulamaiset poikittaiset väliseinät Sisarsolut seulaelementit säilyttäen rakenteensa |
Räni (phloem), sijaitsee juuren, varren, lehtisuonien varrella Sijaitsee aina seulaelementtien varrella (mukana) |
Orgaanisen aineen kuljettaminen lehdistä varteen, juureen, kukkiin Osallistu aktiivisesti orgaanisten aineiden kuljettamiseen seulaputket floem |
| 3. Johtavat verisuoni-kuitukimput | Puun ja niinin kompleksi erillisten säikeiden muodossa ruohossa ja jatkuvana massana puissa | Juuren ja varren keskussylinteri; lehtien ja kukkien suonet | Veden ja mineraalien kuljettaminen puun läpi; partalla - orgaaniset aineet; vahvistaa elimiä yhdistämällä ne yhdeksi kokonaisuudeksi |



Kasvien mekaaninen kudos.


Tämä tyyppi kuuluu monimutkaisiin kudoksiin ja koostuu erilailla erilaistuneista soluista. Itse johtavien elementtien lisäksi kudos sisältää mekaanisia, eritys- ja varastoelementtejä. Johtavat kudokset yhdistävät kaikki kasvin elimet yhtenäinen järjestelmä. Johtavia kudoksia on kahta tyyppiä: ksyleemi ja floemi (kreikaksi ksylon - puu; phloios - kuori, niini). Niissä on sekä rakenteellisia että toiminnallisia eroja.
Ksyleemin johtavat elementit muodostuvat kuolleet solut. Ne kuljettavat vettä ja siihen liuenneita aineita pitkiä matkoja juuresta lehtiin. Floeemin johtavat elementit säilyttävät elävän protoplastin. Ne kuljettavat pitkän matkan fotosynteettisistä lehdistä juureen.
Tyypillisesti ksyleemi ja floemi sijaitsevat kasvirungossa tietyssä järjestyksessä muodostaen kerroksia tai verisuonikimppuja. Rakenteesta riippuen on olemassa useita johtavia nipputyyppejä, jotka ovat ominaisia tietyt ryhmät kasvit. Ksyleemin ja floeemin välissä oleva avoin nippu sisältää kambiumia, joka tarjoaa toissijaista kasvua. Kaksipuolisessa avoimessa nipussa floeemi sijaitsee molemmilla puolilla suhteessa ksylemiin. Suljetut kimput eivät sisällä kambiumia eivätkä siten pysty toissijaiseen paksuuntumiseen. Löydät vielä kaksi samankeskistä nipputyyppiä, joissa joko floemi ympäröi ksylemiä tai ksyleemi ympäröi floemia.
Ksylem (puu). Ksyleemin kehittyminen korkeammissa kasveissa liittyy vedenvaihdon varmistamiseen. Koska vettä poistuu jatkuvasti orvaskeden läpi, kasvin on imeytettävä sama määrä kosteutta ja lisättävä se transpiraatiota suorittaviin elimiin. On syytä ottaa huomioon, että elävän protoplastin läsnäolo vettä johtavissa soluissa hidastaisi kuljetusta suuresti, kuolleet solut osoittautuvat tässä toimivammiksi. Kuolleella solulla ei kuitenkaan ole jäykkyyttä, joten kalvolla on oltava mekaanisia ominaisuuksia. Huomautus: turgesenssi - tilat kasvisolut, kudokset ja elimet, mille? ne muuttuvat elastisiksi johtuen solusisällön paineesta niiden elastisiin kalvoihin. Itse asiassa ksyleemin johtavat elementit koostuvat kuolleista soluista, jotka ovat pitkänomaisia elimen akselia pitkin paksuilla lignified-kuorilla.
Aluksi ksyleemi muodostuu ensisijaisesta meristeemistä - prokambiumista, joka sijaitsee aksiaalisten elinten yläosassa. Ensin erotetaan protoksylemi, sitten metaksylemi. Ksyleemin muodostumista tunnetaan kolmea tyyppiä. Exarch-tyypissä protoksyleemielementit ilmestyvät ensin prokambium-kimpun reunalle, sitten metaksyleemielementit ilmestyvät keskelle. Jos prosessi menee vastakkaiseen suuntaan (eli keskustasta reuna-alueelle), tämä on endarkkinen tyyppi. Mesarkisessa tyypissä ksyleemi muodostuu prokambiaalisen nipun keskelle, jonka jälkeen se kerrostuu sekä keskustaa päin että reunaa kohti.
Juurelle on ominaista eksarkaalinen ksyleemin muodostus, kun taas varrelle on ominaista endarkkinen tyyppi. Matalaorganisoituneissa kasveissa ksyleemin muodostusmenetelmät ovat hyvin erilaisia ja voivat toimia systemaattisina ominaisuuksina.
Jonkin verran? Kasveissa (esimerkiksi yksisirkkaisissa) kaikki prokambiumsolut erilaistuvat johtaviksi kudoksiksi, jotka eivät kykene toissijaiseen paksuuntumiseen. Muissa muodoissa (esimerkiksi puumaisissa) lateraalimeristeemit (kambium) jäävät ksyleemin ja floeemin väliin. Nämä solut pystyvät jakautumaan, uusien ksyleemin ja floemin. Tätä prosessia kutsutaan toissijaiseksi kasvuksi. Monissa kasvaa suhteellisen vakaasti ilmasto-olosuhteet, kasvit, kasvu on jatkuvaa. Vuodenaikojen ilmastomuutoksiin mukautuneissa muodoissa - ajoittain.
Prokambiumsolujen erilaistumisen päävaiheet. Sen soluissa on ohuet kalvot, jotka eivät estä niitä venymästä elimen kasvun aikana. Protoplasti alkaa sitten muodostaa toissijaista kuorta. Mutta tällä prosessilla on erityisiä piirteitä. Toissijainen kuori ei kerrostu jatkuvana kerroksena, joka ei salli solun venymistä, vaan renkaiden tai spiraalin muodossa. Solun pidentäminen ei ole vaikeaa. Nuorissa soluissa heliksin renkaat tai käänteet sijaitsevat lähellä toisiaan. Kypsissä soluissa solut eroavat solujen pidentymisen seurauksena. Kuoren rengas- ja spiraalipaksunteet eivät häiritse kasvua, mutta mekaanisesti ne ovat kuoria huonompia, joissa sekundaarinen paksuuntuminen muodostaa jatkuvan kerroksen. Tässä suhteessa kasvun lakkaamisen jälkeen ksyleemiin muodostuu elementtejä, joissa on jatkuva lignifioitu kuori (metaksyleemi). On huomattava, että toissijainen paksuuntuminen tässä ei ole rengasmainen tai spiraalimainen, vaan pistemäinen, skalarimainen, verkko, jonka solut eivät veny ja kuolevat muutamassa tunnissa. Tämä prosessi tapahtuu koordinoidusti läheisissä soluissa. Ilmestyy sytoplasmassa suuri määrä lysosomit Sitten ne hajoavat ja niiden sisältämät entsyymit tuhoavat protoplastin. Kun poikittaisseinämät tuhoutuvat, ketjussa toistensa yläpuolella olevat solut muodostavat onton suonen. Useimmat koppisiemeniset ja jotkut? pteridofyteilla on verisuonia.
Johtavaa solua, joka ei muodostu seinämässä olevien reikien kautta, kutsutaan trakeidiksi. Vesi liikkuu henkitorven läpi hitaammin kuin alusten läpi. Tosiasia on, että henkitorveissa ensisijainen kuori ei keskeydy missään. Trakeidit kommunikoivat keskenään huokosten kautta. On syytä selventää, että kasveissa huokos on vain syvennys toissijaisessa kuoressa primaarikuoreen asti eikä henkitorven välillä ole läpimeneviä reikiä.
Yleisimmät ovat rajatut huokoset. Niissä soluonteloon päin oleva kanava muodostaa jatkeen - huokoskammion. Useimmat huokoset havupuukasveja primaarisessa kuoressa niillä on paksuus - torus, joka on eräänlainen venttiili ja pystyy säätelemään veden kuljetuksen voimakkuutta. Siirtymällä torus estää veden virtauksen huokosen läpi, mutta sen jälkeen se ei voi enää palata edelliseen asentoonsa suorittamalla kertaluonteisen toimenpiteen.
Huokoset ovat enemmän tai vähemmän pyöreitä, pitkänomaisia kohtisuorassa pitkänomaiseen akseliin nähden (näiden huokosten ryhmä muistuttaa tikkaita; siksi tällaista huokoisuutta kutsutaan portaikoksi). Huokosten kautta kuljetus tapahtuu sekä pituus- että poikittaissuunnassa. Huokosia ei ole vain henkitorveissa, vaan myös yksittäisissä verisuonisoluissa, jotka muodostavat suonen.
Evoluutioteorian näkökulmasta trakeidit edustavat ensimmäistä ja päärakennetta, joka johtaa vettä korkeampien kasvien kehossa. Uskotaan, että suonet syntyivät henkitorveista niiden välisten poikittaisseinien hajoamisen vuoksi. Useimmilla pteridofyyteillä ja -siemenisilla ei ole suonia. Niiden vesi liikkuu henkitorven kautta.
Käynnissä evoluution kehitys alukset ilmestyivät sisään eri ryhmiä kasveja toistuvasti, mutta ne saivat tärkeimmän toiminnallisen merkityksen koppisiemenissä, missä? niitä esiintyy henkitorven mukana. Uskotaan, että kehittyneemmän kuljetusmekanismin hallussapito auttoi heitä paitsi selviytymään, myös saavuttamaan huomattavan erilaisia muotoja.
Ksylem on monimutkainen kudos, joka sisältää vettä johtavien alkuaineiden lisäksi myös muita. Mekaaniset toiminnot suoritetaan libriformisilla kuiduilla (latinaksi liber - bast, forma - muoto). Mekaanisten lisärakenteiden läsnäolo on tärkeää, koska sakeutumisesta huolimatta vettä johtavien elementtien seinämät ovat edelleen liian ohuita. He eivät pysty elättämään suuria massoja yksin. monivuotinen kasvi. Kuidut kehittyivät henkitorveista. Niille on ominaista pienemmät koot, lignified (lignified) kuoret ja kapeat ontelot. Reunuksettomat huokoset löytyvät seinästä. Nämä kuidut eivät johda vettä, vaan niiden päätehtävä on tukea.
Ksyleemissä on myös eläviä soluja. Niiden massa voi olla 25% puun kokonaistilavuudesta. Koska nämä solut ovat muodoltaan pyöreitä, niitä kutsutaan puuparenkyymiksi. Kasvin rungossa parenkyyma sijaitsee kahdella tavalla. Ensimmäisessä tapauksessa solut on järjestetty pystysuorien säikeiden muodossa - tämä on juosteen parenkyymi. Toisessa tapauksessa parenchyma muodostaa vaakasuuntaisia säteitä. Niitä kutsutaan ydinsäteiksi, koska ne yhdistävät ytimen ja aivokuoren. Ydin suorittaa useita toimintoja, mukaan lukien aineiden varastoinnin.
Phloem (bast). Tämä on monimutkainen kudos, koska sen muodostavat erityyppiset solut. Pääjohtavia kennoja kutsutaan seulaelementeiksi. Ksyleemin johtavat elementit muodostuvat kuolleista soluista, kun taas floeemissa ne säilyttävät elävän, vaikkakin voimakkaasti muunnetun protoplastin toimintajakson aikana. Floem suorittaa muovisten aineiden ulosvirtauksen fotosynteettisistä elimistä. Kyky suorittaa eloperäinen aine Kaikkien elävien kasvisolujen hallussa. Ja näin ollen, jos ksylemiä löytyy vain korkeammista kasveista, niin orgaanisten aineiden kuljetus solujen välillä tapahtuu myös alemmissa kasveissa.
Ksyleemi ja floeemi kehittyvät apikaalisista meristeemeistä. Ensimmäisessä vaiheessa protofloemi muodostuu prokambiaaliseen johtoon. Ympäröivien kudosten kasvaessa se venyy, ja kun kasvu on päättynyt, protofloeemin sijaan muodostuu metafloeemi.
Korkeampien kasvien eri ryhmissä löytyy kahden tyyppisiä seulaelementtejä. Pteridofyyteissä ja siemenissä sitä edustavat seulasolut. Kennojen seulakentät ovat hajallaan sivuseiniä pitkin. Protoplasti säilyttää jonkin verran tuhoutuneen ytimen.
Koppisiemenissä seulaelementtejä kutsutaan seulaputkiksi. Ne kommunikoivat keskenään seulalevyjen kautta. Kypsistä soluista puuttuu ytimiä. Seulaputken vieressä on kuitenkin seurasolu, joka muodostuu yhdessä seulaputken kanssa yhteisen emosolun mitoottisen jakautumisen seurauksena (kuva 38). Seurasolulla on tiheämpi sytoplasma, jossa on suuri määrä aktiivisia mitokondrioita, sekä täysin toimiva ydin, valtava määrä plasmodesmataa (kymmenen kertaa enemmän kuin muut solut). Seurasolut vaikuttavat tumaputkiseulasolujen toiminnalliseen aktiivisuuteen.
Kypsien seulasolujen rakenteessa on joitain erityispiirteitä. Vakuolia ei ole, ja siksi sytoplasma nesteytyy suuresti. Ydin voi olla poissa (koppisiemenissä) tai ryppyisessä, toiminnallisesti inaktiivisessa tilassa. Ribosomit ja Golgi-kompleksi puuttuvat myös, mutta endoplasminen retikulumi on hyvin kehittynyt, joka ei vain tunkeudu sytoplasmaan, vaan myös kulkeutuu naapurisoluihin seulakenttien huokosten kautta. Hyvin kehittyneitä mitokondrioita ja plastideja löytyy runsaasti.
Solujen välillä aineet kuljetetaan solukalvoissa olevien reikien kautta. Tällaisia aukkoja kutsutaan huokosiksi, mutta toisin kuin henkitorven huokoset, ne ovat läpi. Oletetaan, että ne edustavat suuresti laajentuneita plasmodesmataa seinillä, mikä? kalloosipolysakkaridi kerrostuu. Huokoset on järjestetty ryhmiin muodostaen seulakenttiä. Primitiivisissä muodoissa seulakentät ovat satunnaisesti hajallaan kuoren koko pinnalle; edistyneemmissä koppisiemenissä ne sijaitsevat vierekkäisten solujen päissä vierekkäin muodostaen seulalevyn. Jos siinä on yksi seulakenttä, sitä kutsutaan yksinkertaiseksi, jos niitä on useita, sitä kutsutaan kompleksiksi.
Liuosten liikkumisnopeus seulaelementtien läpi on jopa 150 cm tunnissa. Tämä on tuhat kertaa nopeampi kuin vapaan diffuusion nopeus. Todennäköisesti tapahtuu aktiivista kuljetusta, ja lukuisat seulaelementtien mitokondriot ja seurasolut tarjoavat tähän tarvittavan ATP:n.
Floem-seulaelementtien aktiivisuuden kesto riippuu lateraalisten meristeemien läsnäolosta. Jos niitä on, seulaelementit toimivat koko kasvin käyttöiän ajan.
Seulaelementtien ja seurasolujen lisäksi floemi sisältää niinikuituja, sklereideja ja parenkyymiä.
Johtavia kankaita
Tämä tyyppi kuuluu monimutkainen kudoksia, koostuu erilaistuneista soluista. Itse johtavien elementtien lisäksi kudos sisältää mekaanisia, eritys- ja varastoelementtejä (kuva 26). Johtavat kudokset yhdistävät kaikki kasvin elimet yhdeksi järjestelmäksi. Johtavia kudoksia on kahta tyyppiä: ksylem Ja floem(kreikaksi ksylon - puu; phloios - kuori, niini). Niissä on sekä rakenteellisia että toiminnallisia eroja.
Ksyleemin johtavat elementit muodostuvat kuolleista soluista. Ne kuljettavat vettä ja siihen liuenneita aineita pitkiä matkoja juuresta lehtiin. Floeemin johtavat elementit säilyttävät elävän protoplastin. Ne kuljettavat pitkän matkan fotosynteettisistä lehdistä juureen.
Tyypillisesti ksyleemi ja floeemi sijaitsevat kasvirungossa tietyssä järjestyksessä muodostaen kerroksia tai johtavia nippuja. Rakenteesta riippuen on olemassa useita erityyppisiä verisuonikimppuja, jotka ovat ominaisia tietyille kasviryhmille. Vakuudeksi avoimessa nipussa Ksyleemin ja floeemin välissä on kambium, joka saa aikaan sekundaarikasvun (kuvat 27-A, 28). Kaksipuolisessa avoimessa nipussa floemi sijaitsee suhteessa ksylemiin molemmilla puolilla (kuvat 27-B, 29). Suljetut niput eivät sisällä kambiumia eivätkä siten pysty toissijaiseen paksuuntumiseen (kuvat 27-B, 27-D, 30,31). Kaksi muuta tyyppiä löytyy samankeskiset palkit, jossa joko floeemi ympäröi ksylemiä (Kuva 27-D, 32) tai ksyleemi ympäröi floeemia (Kuva 27-E).
Ksylem (puu). Ksyleemin kehittyminen korkeammissa kasveissa liittyy vedenvaihdon varmistamiseen. Koska vettä poistuu jatkuvasti orvaskeden läpi, kasvin on imeytettävä sama määrä kosteutta ja lisättävä se transpiraatiota suorittaviin elimiin. On syytä ottaa huomioon, että elävän protoplastin läsnäolo vettä johtavissa soluissa hidastaisi kuljetusta suuresti, kuolleet solut osoittautuvat tässä toimivammiksi. Kuolleella solulla ei kuitenkaan ole turvotus Siksi kuorella on oltava mekaanisia ominaisuuksia. Huomautus: turgesenssi - kasvisolujen, kudosten ja elinten tila, jossa ne muuttuvat elastisiksi johtuen solusisällön paineesta niiden elastisiin kalvoihin. Itse asiassa ksyleemin johtavat elementit koostuvat kuolleista soluista, jotka ovat pitkänomaisia elimen akselia pitkin paksuilla lignified-kuorilla.
Aluksi ksyleemi muodostuu ensisijaisesta meristeemistä - prokambiumista, joka sijaitsee aksiaalisten elinten yläosassa. Ensin erotettu protoksilemi, sitten metaksylemi. Ksyleemin muodostumista tunnetaan kolmea tyyppiä. klo exarch Tässä tyypissä protoksyleemielementit ilmestyvät ensin prokambium-kimpun reunalle, sitten metaksyleemielementit ilmestyvät keskelle. Jos prosessi menee päinvastaiseen suuntaan (eli keskustasta reuna-alueelle), niin tämä endarkkinen tyyppi. klo mesarkaalinen tyyppi ksyleemi käynnistyy prokambiaalisen nipun keskeltä, minkä jälkeen se laskeutuu sekä keskustaan että reunaan päin.
Juurelle on ominaista eksarkaalinen ksyleemin muodostus, kun taas varrelle on ominaista endarkkinen tyyppi. Matalaorganisoituneissa kasveissa ksyleemin muodostusmenetelmät ovat hyvin erilaisia ja voivat toimia systemaattisina ominaisuuksina.
Joissakin kasveissa (esimerkiksi yksisirkkaisissa) kaikki prokambiumsolut erilaistuvat johtaviksi kudoksiksi, jotka eivät pysty sekundaariseen paksuuntumiseen. Muissa muodoissa (esimerkiksi puumaisissa) lateraalimeristeemit (kambium) jäävät ksyleemin ja floeemin väliin. Nämä solut pystyvät jakautumaan, uusien ksyleemin ja floemin. Tätä prosessia kutsutaan toissijainen kasvu. Monet suhteellisen vakaissa ilmasto-oloissa kasvavat kasvit kasvavat jatkuvasti. Vuodenaikojen ilmastomuutoksiin mukautuneissa muodoissa - ajoittain. Tämän seurauksena muodostuu hyvin määritellyt vuosikasvurenkaat.
Prokambiumsolujen erilaistumisen päävaiheet. Sen soluissa on ohuet kalvot, jotka eivät estä niitä venymästä elimen kasvun aikana. Protoplasti alkaa sitten muodostaa toissijaista kuorta. Mutta tällä prosessilla on erityisiä piirteitä. Toissijainen kuori ei kerrostu jatkuvana kerroksena, joka ei salli solun venymistä, vaan renkaiden tai spiraalin muodossa. Solun pidentäminen ei ole vaikeaa. Nuorissa soluissa heliksin renkaat tai käänteet sijaitsevat lähellä toisiaan. Kypsissä soluissa solut eroavat solujen venytyksen seurauksena (kuva 33). Kuoren rengas- ja spiraalipaksunteet eivät häiritse kasvua, mutta mekaanisesti ne ovat kuoria huonompia, joissa sekundaarinen paksuuntuminen muodostaa jatkuvan kerroksen. Siksi kasvun pysähtymisen jälkeen ksyleemiin muodostuu elementtejä, joissa on jatkuva lignoitunut kuori ( metaksylemi). On huomattava, että toissijainen paksuuntuminen ei ole renkaan muotoinen tai spiraalimainen, vaan pistemäinen, skalarimainen ja retikulaarinen (kuva 34). Sen solut eivät pysty venymään eivätkä kuole muutamassa tunnissa. Tämä prosessi tapahtuu koordinoidusti läheisissä soluissa. Sytoplasmassa esiintyy suuri määrä lysosomeja. Sitten ne hajoavat ja niiden sisältämät entsyymit tuhoavat protoplastin. Kun poikittaisseinämät tuhoutuvat, ketjussa toistensa yläpuolella olevat solut muodostavat onton suonen (kuva 35). Useimmissa koppisiemenissä ja joissakin pteridofyytissä on suonia.
Johtavaa solua, joka ei muodostu seinämässä olevien reikien kautta, kutsutaan trakeidi. Vesi liikkuu henkitorven läpi hitaammin kuin alusten läpi. Tosiasia on, että henkitorveissa ensisijainen kuori ei keskeydy missään. Trakeidit kommunikoivat keskenään por. On syytä selventää, että kasveissa huokos on vain syvennys toissijaisessa kuoressa primaarikuoreen asti eikä henkitorven välillä ole läpimeneviä reikiä.
Useimmiten löytyy rajattuja huokosia (kuva 35-1). Niissä soluonteloon päin oleva kanava muodostaa jatkeen - huokoskammio. Useimpien havupuukasvien huokoset ensisijaisessa kuoressa ovat paksuuntuneet - torus, joka on eräänlainen venttiili ja pystyy säätelemään veden kuljetuksen voimakkuutta. Siirtymällä torus estää veden virtauksen huokosen läpi, mutta sen jälkeen se ei voi enää palata edelliseen asentoonsa suorittamalla kertaluonteisen toimenpiteen.
Huokoset ovat enemmän tai vähemmän pyöreitä, pitkänomaisia kohtisuorassa pitkänomaiseen akseliin nähden (ryhmä näistä huokosista muistuttaa tikkaita, minkä vuoksi tällaista huokoisuutta kutsutaan portaikoksi). Huokosten kautta kuljetus tapahtuu sekä pituus- että poikittaissuunnassa. Huokosia ei ole vain henkitorveissa, vaan myös yksittäisissä verisuonisoluissa, jotka muodostavat suonen.
Evoluutioteorian näkökulmasta trakeidit edustavat ensimmäistä ja päärakennetta, joka johtaa vettä korkeampien kasvien kehossa. Uskotaan, että suonet syntyivät henkitorveista niiden välisten poikittaisseinien hajoamisen vuoksi (kuva 36). Useimmilla pteridofyyteillä ja -siemenisilla ei ole suonia. Niiden vesi liikkuu henkitorven kautta.
Evoluutiokehitysprosessissa suonia esiintyi eri kasviryhmissä useammin kuin kerran, mutta ne saivat tärkeimmän toiminnallisen merkityksen koppisiemenissä, joissa niitä esiintyy henkitorven mukana. Uskotaan, että kehittyneemmän kuljetusmekanismin hallussapito auttoi heitä paitsi selviytymään, myös saavuttamaan huomattavan erilaisia muotoja.
Ksylem on monimutkainen kudos, joka sisältää vettä johtavien alkuaineiden lisäksi myös muita. Mekaaniset toiminnot suoritetaan libriformisilla kuiduilla ( lat. liber - bast, forma - muoto). Mekaanisten lisärakenteiden läsnäolo on tärkeää, koska sakeutumisesta huolimatta vettä johtavien elementtien seinämät ovat edelleen liian ohuita. He eivät pysty itsenäisesti tukemaan suurta monivuotisen kasvin massaa. Kuidut kehittyivät henkitorveista. Niille on ominaista pienemmät koot, lignified (lignified) kuoret ja kapeat ontelot. Reunuksettomat huokoset löytyvät seinästä. Nämä kuidut eivät johda vettä, vaan niiden päätehtävä on tukea.
Ksyleemissä on myös eläviä soluja. Niiden massa voi olla 25% puun kokonaistilavuudesta. Koska nämä solut ovat muodoltaan pyöreitä, niitä kutsutaan puuparenkyymiksi. Kasvin rungossa parenkyyma sijaitsee kahdella tavalla. Ensimmäisessä tapauksessa solut on järjestetty pystysuorien säikeiden muodossa - tämä on säikeen parenkyymi. Toisessa tapauksessa parenchyma muodostaa vaakasuuntaisia säteitä. Niitä kutsutaan medullaariset säteet, koska ne yhdistävät ytimen ja kuoren. Ydin suorittaa useita toimintoja, mukaan lukien aineiden varastoinnin.
Phloem (bast). Tämä on monimutkainen kudos, koska sen muodostavat erityyppiset solut. Pääjohtavia soluja kutsutaan seulaelementit(Kuva 37). Ksyleemin johtavat elementit muodostuvat kuolleista soluista, kun taas floeemissa ne säilyttävät elävän, vaikkakin voimakkaasti muunnetun protoplastin toimintajakson aikana. Floem suorittaa muovisten aineiden ulosvirtauksen fotosynteettisistä elimistä. Kaikilla elävillä kasvisoluilla on kyky johtaa orgaanisia aineita. Ja näin ollen, jos ksylemiä löytyy vain korkeammista kasveista, niin orgaanisten aineiden kuljetus solujen välillä tapahtuu myös alemmissa kasveissa.
Ksyleemi ja floeemi kehittyvät apikaalisista meristeemeistä. Ensimmäisessä vaiheessa a protofloem. Kun ympäröivät kudokset kasvavat, se venyy, ja kun kasvu on valmis, sen sijaan muodostuu protofloeemi metafloeemi.
Korkeampien kasvien eri ryhmissä löytyy kahden tyyppisiä seulaelementtejä. Se on edustettuna pteridofyyteissä ja kimnosiemenissä seulasolut. Kennojen seulakentät ovat hajallaan sivuseiniä pitkin. Protoplasti säilyttää jonkin verran tuhoutuneen ytimen.
Koppisiemenissä kutsutaan seulaelementtejä seulaputket. Ne kommunikoivat keskenään seulalevyjen kautta. Kypsistä soluista puuttuu ytimiä. Seulaputken vieressä on kuitenkin seuralainen solu, muodostuu yhdessä seulaputken kanssa yhteisen emosolun mitoottisen jakautumisen seurauksena (kuva 38). Seurasolulla on tiheämpi sytoplasma, jossa on suuri määrä aktiivisia mitokondrioita, sekä täysin toimiva ydin, valtava määrä plasmodesmataa (kymmenen kertaa enemmän kuin muut solut). Seurasolut vaikuttavat tumaputkiseulasolujen toiminnalliseen aktiivisuuteen.
Kypsien seulasolujen rakenteessa on joitain erityispiirteitä. Vakuolia ei ole, joten sytoplasma nesteytyy suuresti. Ydin voi olla poissa (koppisiemenissä) tai ryppyisessä, toiminnallisesti inaktiivisessa tilassa. Ribosomit ja Golgi-kompleksi puuttuvat myös, mutta endoplasminen retikulumi on hyvin kehittynyt, joka ei vain tunkeudu sytoplasmaan, vaan myös kulkeutuu naapurisoluihin seulakenttien huokosten kautta. Hyvin kehittyneitä mitokondrioita ja plastideja löytyy runsaasti.
Solujen välillä aineet kuljetetaan solukalvoissa olevien reikien kautta. Tällaisia aukkoja kutsutaan huokosiksi, mutta toisin kuin henkitorven huokoset, ne ovat läpi. Oletetaan, että ne ovat voimakkaasti laajentuneita plasmodesmaatteja, joiden seinämille on kerrostunut kalloosipolysakkaridi. Huokoset on järjestetty ryhmiin muodostaen seulakentät. Primitiivisissä muodoissa seulakentät ovat satunnaisesti hajallaan kuoren koko pinnalle; edistyneemmissä koppisiemenissä ne sijaitsevat vierekkäisten solujen päissä vierekkäin muodostaen seulalevy(Kuva 39). Jos siinä on yksi seulakenttä, sitä kutsutaan yksinkertaiseksi, jos niitä on useita, sitä kutsutaan kompleksiksi.
Liuosten liikkumisnopeus seulaelementtien läpi on jopa 150 cm? tunnin. Tämä on tuhat kertaa nopeampi kuin vapaan diffuusion nopeus. Todennäköisesti tapahtuu aktiivista kuljetusta, ja lukuisat seulaelementtien mitokondriot ja seurasolut tarjoavat tähän tarvittavan ATP:n.
Floem-seulaelementtien aktiivisuuden kesto riippuu lateraalisten meristeemien läsnäolosta. Jos niitä on, seulaelementit toimivat koko kasvin käyttöiän ajan.
Seulaelementtien ja seurasolujen lisäksi floemi sisältää niinikuituja, sklereideja ja parenkyymiä.
Johtavat kudokset ovat monimutkaisia, koska ne koostuvat useista solutyypeistä, niiden rakenteessa on pitkänomainen (putkimainen) muoto ja useat huokoset läpäisevät sen. Reikien läsnäolo päätyosissa (ala- tai yläosissa) mahdollistaa pystysuoran kuljetuksen ja sivupintojen huokoset helpottavat veden virtausta säteittäiseen suuntaan. Johtaviin kudoksiin kuuluvat ksyleemi ja floeemi. Niitä löytyy vain saniaisten kaltaisissa ja siemeniä kantavissa kasveissa. Johtava kudos sisältää sekä kuolleita että eläviä soluja
Ksylem (puu)- Tämä on kuollutta kudosta. Sisältää perus rakenneosat(henkitorvi ja henkitorvi), puuparenkyymi ja puukuidut. Se suorittaa kasvessa sekä tukevaa että johtavaa tehtävää - vesi ja mineraalisuolat kulkevat kasvessa sen läpi ylös.
Trakeidit
– kuolleet yksittäiset karan muotoiset solut. Seinät ovat paksuuntuneet huomattavasti ligniinin laskeuman vuoksi. Henkitorvien erityispiirre on rajattujen huokosten läsnäolo niiden seinissä. Niiden päät menevät päällekkäin, mikä antaa kasville tarvittavan lujuuden. Vesi liikkuu henkitorven tyhjien onteloiden läpi ilman, että se kohtaa matkalla mitään häiriöitä solusisällön muodossa; se siirtyy henkitorvesta toiseen huokosten kautta.
Koppisiemenissä trakeidit ovat kehittyneet verisuonet (henkitorvi). Nämä ovat erittäin pitkiä putkia, jotka muodostuvat useiden solujen "liitoksen" seurauksena; päätyväliseinien jäänteet ovat edelleen säilyneet astioissa reunusten muodossa. Alusten koot vaihtelevat useista sentteistä useisiin metriin. Ensimmäisessä muodostuvassa protoksyleemissä ligniini kerääntyy renkaisiin tai spiraaliin. Tämä antaa suonen jatkaa venymistä sen kasvaessa. Metaksyleemiastioissa ligniini keskittyy tiheämmin - tämä on ihanteellinen "vesiputki", joka toimii pitkiä matkoja.
?1.
Miten henkitorvet eroavat henkitorveista? (Vastaus artikkelin lopussa)
?2
. Miten trakeidit eroavat kuiduista?
?3
. Mitä yhteistä on floemilla ja ksyleemillä?
?4.
Miten seulaputket eroavat henkitorvesta?
Ksyleemin parenkyymisolut muodostavat omituisia säteitä, jotka yhdistävät ytimen kuoreen. Ne johtavat vettä säteittäiseen suuntaan, varastoivat ravinteita. Uudet ksyleemisuonet kehittyvät muista parenkyymisoluista. Lopuksi puukuidut ovat samanlaisia kuin trakeidit, mutta toisin kuin niillä on hyvin pieni sisäontelo, joten ne eivät johda vettä, vaan antavat lisälujuutta. Niissä on myös yksinkertaiset huokoset, eivät rajattuja.
Phloem (pesä)- Tämä elävää kudosta, joka on osa kasvin kuorta, suorittaa alaspäin suuntautuva virta vettä ja siihen liuenneita assimilaatiotuotteita. Phloem muodostuu viiden tyyppisestä rakenteesta: seulaputket, seurasolut, niiniparenkyymi, niinikuidut ja sklereidit.
Näiden rakenteiden perusta on seulaputket
, muodostuu useiden seulakennojen yhdistämisen seurauksena. Niiden seinämät ovat ohuita, selluloosaa, ytimet kuolevat kypsymisen jälkeen ja sytoplasma puristuu seinämiä vasten, mikä vapauttaa tien orgaanisille aineille. Seulaputkien kennojen päätyseinät peittyvät vähitellen huokosiin ja alkavat muistuttaa seulaa - nämä ovat seulalevyjä. Elintoimintojensa varmistamiseksi seurasolut sijaitsevat lähellä, niiden sytoplasma on aktiivinen ja niiden tumat ovat suuria.
?5
. Miksi luulet, että kun seulasolut kypsyvät, niiden tumat kuolevat?
VASTAUKSIA
?1.
Henkitorvet ovat monisoluisia rakenteita, eikä niillä ole päätyseinämiä, mutta henkitorvi on yksisoluinen, niillä on päätyseinät ja reunustetut huokoset.
?2
. Henkitorveilla on rajalliset huokoset ja hyvin rajattu ontelo, kun taas kuiduilla on hyvin pieni ontelo ja yksinkertaiset huokoset. Ne eroavat myös toiminnaltaan, henkitorveilla on kuljetustehtävä (johtava) ja kuiduilla mekaaninen rooli.
?3.
Floem ja ksyleemi ovat molemmat johtavia kudoksia; niiden rakenteet ovat muodoltaan putkimaisia ja sisältävät parenkyymisoluja ja mekaanisia kudoksia.
?4.
Seulaputket koostuvat elävistä soluista, niiden seinämät ovat selluloosaa, ne kuljettavat orgaanisia aineita alaspäin ja henkitorven muodostavat kuolleet solut; niiden seinämät ovat voimakkaasti paksuntuneet ligniinillä, ne kuljettavat vettä ja mineraaleja ylöspäin.
?5.
Kuljetus alaspäin tapahtuu seulasoluja pitkin ja ainevirran mukana kuljettamat ytimet peittäisivät merkittävän osan seulakentästä, mikä johtaisi prosessin tehokkuuden laskuun.
Johtava kudos koostuu elävistä tai kuolleista pitkänomaisista soluista, jotka näyttävät putkilta.
Kasvien varret ja lehdet sisältävät nippuja johtavaa kudosta. Johtava kudos sisältää suonia ja seulaputkia.
Alukset- peräkkäin kytketyt kuolleet ontot solut, niiden väliset poikittaisväliseinät katoavat. Suonten kautta juurista liuenneet vesi ja mineraalit pääsevät varteen ja lehtiin.
Seulaputket - pitkänomaiset, ytimettömät elävät solut, jotka on kytketty sarjaan toisiinsa. Niiden kautta lehdistä (jossa ne muodostuivat) orgaaniset aineet siirtyvät kasvin muihin elimiin.
Johtava kangas varmistaa veden ja siihen liuenneiden mineraalien kulkeutumisen.
Tämä kudos muodostaa kaksi kuljetusjärjestelmää:
- ylöspäin(juurista lehtiin);
- alaspäin(lehdistä kaikkiin muihin kasvin osiin).
Nouseva kuljetusjärjestelmä koostuu henkitorveista ja suonista (ksyleemi tai puu), ja suonet ovat kehittyneempiä johtimia kuin trakeidit.
Laskevissa järjestelmissä fotosynteesituotteita sisältävä vesivirta kulkee seulaputkien (floem tai floem) läpi.
Ksyleemi ja floemi muodostavat verisuonikuituisia nippuja - kasvin "verenkiertojärjestelmän", joka tunkeutuu sen kokonaan ja yhdistää sen yhdeksi kokonaisuudeksi.
Tutkijat uskovat, että kudosten syntyminen liittyy maapallon historiassa kasvien ilmaantumiseen maalle. Kun osa kasvista joutui ilmaan ja toinen osa (juuri) maaperään, tuli tarpeelliseksi kuljettaa vettä ja mineraalisuoloja juurista lehtiin ja orgaanisia aineita lehdistä juuriin. Siis evoluution aikana kasvisto Syntyi kahden tyyppisiä johtavia kankaita - puu ja niini.
Puun läpi (trakeidien ja suonien kautta) vesi liuenneena mineraalit nousee juurista lehtiin - tämä on vettä johtava tai nouseva virta. Floemin kautta (seulaputkien läpi) vihreissä lehdissä muodostuneet orgaaniset aineet virtaavat kasvin juuriin ja muihin elimiin - tämä on alaspäin suuntautuva virta.
Koulutuskangas
Kasvatuskudosta löytyy kaikista kasvin kasvavista osista. Kasvatuskudos koostuu soluista, jotka pystyvät jakautumaan koko kasvin elinkaaren ajan. Täällä olevat solut valehtelevat hyvin nopeasti toisilleen. Jakautuessaan ne muodostavat monia uusia soluja, mikä varmistaa kasvin pituuden ja paksuuden. Koulukudosten jakautumisen aikana ilmaantuvat solut muunnetaan sitten muiden kasvikudosten soluiksi.

Tämä primaarinen kudos, josta kaikki muut kasvikudokset muodostuvat. Se koostuu erityisistä soluista, jotka pystyvät jakautumaan useita kertoja. Nämä solut muodostavat minkä tahansa kasvin alkion.
Tämä kudos säilyy aikuisessa kasvissa. Se sijaitsee:
- juurijärjestelmän alaosassa ja varsien yläosissa (varmistaa kasvien kasvun korkeudessa ja juurijärjestelmän kehityksen) - apikaalinen koulutuskudos;
- varren sisällä (varmistaa kasvin leveyden ja paksuuntumisen) - sivuttaiskasvatuskudos.
Toisin kuin muut kudokset, sytoplasma koulutus kangas paksumpi ja tiheämpi. Solussa on hyvin kehittyneet organellit, jotka tarjoavat proteiinisynteesiä. Ytimelle on ominaista suuri koko. Ytimen ja sytoplasman massa pidetään vakiona. Ytimen laajentuminen merkitsee solunjakautumisprosessin alkamista, mikä tapahtuu mitoosin kautta kasvien vegetatiivisille osille ja meioosin kautta sporogeenisille meristeemeille.





