Mitohondriji sadrže kružnu DNK. Istraživanje mitohondrijske DNA. Svojstva i funkcije DNK
Zašto mitohondriji trebaju vlastitu DNK? Premda zašto simbionti ne bi imali u sebi vlastitu DNK koja na licu mjesta proizvodi sve što im je potrebno? Zašto onda prenositi dio mitohondrijske DNA u jezgru stanice, stvarajući potrebu za transportom genskih proizvoda u mitohondrije? Zašto se mitohondriji prenose samo od jednog roditelja? Kako se mitohondriji dobiveni od majke slažu s genomom stanice, sačinjenom od DNK majke i oca? Što više ljudi saznaje o mitohondrijima, postavlja se više pitanja.

Međutim, to se ne odnosi samo na mitohondrije: u bilo kojem području bilo koje znanosti proširenje sfere znanja dovodi samo do povećanja njezine površine u dodiru s nepoznatim, uzrokujući sve više pitanja na koja će odgovori proširiti istu sferu s istim predvidljivim rezultatom.
Dakle, DNK modernih mitohondrija raspoređena je na vrlo čudan način: mali dio gena nalazi se izravno u mitohondrijima na kružnom kromosomu (točnije, u nekoliko kopija istog kromosoma u svakom mitohondriju), a većina nacrta za proizvodnju mitohondrija sastavni dijelovi pohranjeni su u staničnoj jezgri. Stoga se kopiranje ovih gena događa istodobno s kopiranjem genoma cijelog organizma, a proizvodi koje oni proizvode odlaze daleko od citoplazme stanice u mitohondrije. Ipak, to je na mnogo načina prikladno: mitohondriji su oslobođeni potrebe kopiranja svih ovih gena tijekom reprodukcije, čitanja i izgradnje proteina i drugih komponenti, usredotočujući se na svoju glavnu funkciju proizvodnje energije. Zašto onda u mitohondrijima postoji mala DNK za čije održavanje su potrebni svi ti mehanizmi, oslobađajući se od kojih bi mitohondriji mogli baciti još više resursa na glavnu svrhu svog postojanja?

Isprva se pretpostavljalo da je preostala DNA u mitohondrijima atavizam, naslijeđe pro-mitohondrija koje je apsorbirao metanogen, a koji ima potpuni bakterijski genom. Na početku njihove simbioze, unatoč postojanju tih mitohondrijskih gena u jezgri ( m-geni), koje su bile potrebne za održavanje ugodnog okruženja za pro-mitohondrije unutar metanogena (to je detaljno opisano u odjeljku o mitohondrijima), isti geni su pohranjeni u svakoj od mitohondrija. Pro-mitohondriji su na početku svog života kao simbiont izgledali otprilike isto kao i moderna bakterija na dijagramu lijevo od ovog odlomka.
I vrlo sporo, zbog nedostatka potražnje, ti su geni nestali iz mitohondrijskog kromosoma kao posljedica različitih mutacija. No stanična jezgra akumulirala je sve više m-gena koji su iz uništenih simbionta-mitohondrija ušli u citoplazmu i integrirali se u genom eukariotske himere. Čim se novoizgrađeni m-gen počeo čitati, stanični mehanizmi proizvodili su proizvode potrebne za mitohondrije, oslobađajući simbionte od njihovog neovisnog stvaranja. To znači da se mitohondrijski analog gena koji je prešao u jezgru više nije održavao u radnom stanju prirodnom selekcijom i izbrisan je mutacijama na isti način kao i svi prethodni. Stoga bi bilo logično pretpostaviti da će uskoro oni geni koji su još u mitohondrijima prijeći u jezgru, što će dovesti do velike energetske koristi za eukariote: uostalom, glomazni mehanizmi kopiranja, čitanja i ispravljanja DNK mogu se ukloniti iz svakog mitohondrija, i tako sve što vam je potrebno za stvaranje bjelančevina.

Došavši do ovog zaključka, znanstvenici su izračunali koliko je vremena trebalo svim genima da migriraju kroz prirodni pomak od mitohondrija do jezgre. I pokazalo se da je to razdoblje odavno prošlo. U vrijeme pojave eukariotske stanice, mitohondriji su imali zajednički bakterijski genom od nekoliko tisuća gena (znanstvenici utvrđuju što je to bio genom proučavajući m-gene prenesene u jezgru u različitim organizmima), a sada i mitohondrije svih vrsta eukariota izgubili su od 95 do 99,9% svojih gena. Nitko nije ostavio više od stotinu gena u mitohondrijima, ali niti nitko nema mitohondrije bez gena. Da je slučajnost imala ključnu ulogu u ovom procesu, tada bi barem nekoliko vrsta već prošlo putem prijenosa gena do jezgre do kraja. No, to se nije dogodilo, a mitohondriji različitih vrsta koje su u ovom trenutku proučavane, gubeći gene neovisno jedna o drugoj, zadržale su isti njihov skup, što izravno ukazuje na potrebu prisutnosti upravo tih gena u mitohondrijima.

Štoviše, i druge organele stanica koje proizvode energiju, kloroplasti, također imaju vlastitu DNK, a na isti način, kloroplasti različitih vrsta evoluirali su paralelno i neovisno, svaki s istim skupom gena.
To znači da su sve te značajne neugodnosti u održavanju vlastitog genoma u svakoj mitohondriji stanice (a u prosjeku jedna stanica sadrži nekoliko stotina!) I glomazni aparati za njezino kopiranje-ispravljanje-prevođenje (glavni, ali ne svi!) Su nadjačani po nečemu.
I u ovom trenutku postoji dosljedna teorija o tom "nečemu": sposobnost stvaranja određenih detalja mitohondrija izravno unutar nužno je regulirati brzinu disanja i prilagoditi procese koji se događaju u mitohondrijima svake minute mijenjanja potreba cijeli organizam.

Zamislite da jednoj od stotina mitohondrija stanice iznenada nedostaju elementi respiratornog lanca (vidi detalje o tome u) ili u njemu nema dovoljno ATP sintaza. Ispostavilo se da je ili preopterećen hranom i kisikom i ne može ih obraditi dovoljno brzo, ili mu međumembranski prostor pršti protonima koji nemaju kamo otići - potpuna katastrofa općenito. Naravno, sva ta odstupanja od idealne životne situacije pokreću više signala čiji je cilj izravnati kotrljanje broda koji tone.
Ovi signali pokreću proizvodnju točno onih dijelova koji trenutno nedostaju u mitohondrijima, aktivirajući čitanje gena pomoću kojih se proteini grade. Čim mitohondrija ima dovoljno komponenti respiratornog lanca ili ATP -aza, "kotrljanje će se izjednačiti", signali o potrebi izgradnje novih dijelova prestat će dolaziti, a geni će se ponovno isključiti. Ovo je jedan od iznenađujuće elegantnih u svojoj jednostavnosti neophodnih mehanizama stanične samoregulacije, najmanji njegov poremećaj dovodi do ozbiljne bolesti ili čak do neodrživosti organizma.

Pokušajmo logički odrediti gdje bi se trebali nalaziti geni potrebni za reagiranje na ovaj signal stradanja. Zamislite situaciju da se ti geni nalaze u jezgri stanice koja sadrži par stotina mitohondrija. U jednoj od mitohondrija, na primjer, nedostatak NADH dehidrogenaza: prvi enzim u respiratornom lancu, čija je uloga oduzeti dva elektrona iz molekule NADH, prenijeti ih u sljedeći enzim i pumpati 2-4 protona kroz membranu.
Zapravo, takvi nedostaci bilo kojeg enzima javljaju se prilično često, jer povremeno zakazuju, količina konzumirane hrane se stalno mijenja, potrebe stanice za ATP -om također skaču nakon skokova ili filcanja tijela koje sadrži tu stanicu. Stoga je situacija vrlo tipična. I tako mitohondrije emitiraju signal: "Moramo izgraditi više NADH dehidrogenaze!" Prema staničnim standardima, vrijeme tranzita ovog signala je vrlo značajno, a zapravo je također potrebno povući izgrađenu glasničku RNA iz jezgre u citoplazmu, iz nje stvoriti proteine, poslati ih u mitohondrije ...
I tu nastaje problem koji je mnogo značajniji od gubitka dodatnog vremena: pri stvaranju specijaliziranih proteina mitohondrija, oni su označeni signalom "dostavi u mitohondrije", ali koji? Nepoznato. Stoga proteini koji im nisu potrebni počinju ulaziti u svaki od par stotina mitohondrija. Stanica troši resurse na njihovu proizvodnju i isporuku, mitohondriji su ispunjeni dodatnim respiratornim lancima (što dovodi do neučinkovitosti respiratornih procesa), a jedini mitohondriji kojima su potrebni ti proteini ne primaju ih u dovoljnim količinama, jer ih u najboljem slučaju dobivaju , stoti dio proizvedenog. Stoga ona nastavlja slati signale nevolje, a kaos se nastavlja. Čak je i iz ovog lirskog i površnog opisa onoga što se događa jasno da takva stanica nije održiva. I da postoje geni koji se moraju čitati i prevesti izravno u mitohondrije kako bi se regulirali procesi koji se u njemu odvijaju, a ne oslanjati se na plan proizvodnje noktiju koji je pokrenula strana jezgre ... odnosno proteini dišni lanac za sve mitohondrije odjednom.

Nakon što smo provjerili što točno proizvode različiti organizmi koji ostaju u mitohondrijima (i stoga premještaju m-gene u jezgru neovisno jedan o drugom), otkrili smo da su to elementi za izgradnju respiratornih lanaca i ATPaze, kao i ribosomi (to jest, glavni dio uređaja za emitiranje).
Više o ovome (i ne samo) možete pročitati iz Lane in "Energija, seks, samoubojstvo: mitohondrije i smisao života"... Pa, jednostavno možete usporediti dijagram mitohondrijske DNA, gdje su kodirani proizvodi dekodirani (desno od ovog odlomka), s dijagramom respiratornog lanca (gore), tako da postane jasno što se točno proizvodi u mitohondrijima. Naravno, nisu svi proteini umetnuti u ovaj lanac proizvedeni lokalno; neki od njih izgrađeni su u citoplazmi stanice. No glavna "sidra" za koja se drže ostali dijelovi stvorena su unutar mitohondrija. To vam omogućuje da proizvedete točno onoliko enzima koliko vam je potrebno i točno tamo gdje su potrebni.
Kako su mitohondriji povezani sa spolom i kako različiti genomi koegzistiraju u jednoj stanici, napisat ću u jednom od sljedećih poglavlja ovog retka.
Uvod
Četvrt stoljeća prošlo je od otkrića molekula DNA u mitohondrijima prije nego što su se za njih zainteresirali molekularni biolozi i citolozi, kao i genetičari, evolucionisti, kao i paleontolozi i forenzičari. Tako velik interes izazvao je rad A. Wilson s Kalifornijskog sveučilišta. Godine 1987. objavio je rezultate usporedne analize DNK mitohondrija uzete od 147 predstavnika različitih etničkih skupina svih ljudskih rasa nastanjenih na pet kontinenata. Vrstom, mjestom i brojem pojedinačnih mutacija utvrđeno je da je sva mitohondrijska DNA nastala divergencijom iz jedne sekvence nukleotida predaka. U pseudoznanstvenom tisku ovaj je zaključak protumačen na krajnje pojednostavljen način - cijelo je čovječanstvo potjecalo od jedne žene, po imenu mitohondrijska Eva (budući da i kćeri i sinovi primaju mitohondrije samo od svoje majke), koja je živjela u sjeveroistočnoj Africi oko 200 tisuća godine ... Nakon još 10 godina bilo je moguće dešifrirati fragment mitohondrijske DNA izolirane iz neandertalskih osa, te procijeniti životni vijek posljednjeg zajedničkog pretka čovjeka i neandertalca prije 500 tisuća godina.
Danas se ljudska mitohondrijska genetika intenzivno razvija kako u populaciji tako i u medicinskom aspektu. Uspostavljena je veza između niza teških nasljednih bolesti i defekata u mitohondrijskoj DNA. Genetske promjene povezane sa starenjem najizraženije su u mitohondrijima. Što je mitohondrijski genom, koji se po veličini, obliku i genetskom kapacitetu razlikuje kod ljudi i drugih životinja od biljaka, gljiva i protozoa? Koja je uloga, kako to funkcionira i kako je mitohondrijski genom nastao u različitim svojtama općenito, a posebno kod ljudi? O tome će biti riječi u mom "najmanjem i najskromnijem" eseju.
Osim DNK, mitohondrijski matriks sadrži vlastite ribosome, koji se po mnogim karakteristikama razlikuju od eukariotskih ribosoma koji se nalaze na membranama endoplazmatskog retikuluma. Međutim, na ribosomima mitohondrija ne nastaje više od 5% svih proteina koji čine njihov sastav. Većina proteina koji čine strukturne i funkcionalne komponente mitohondrija kodirani su nuklearnim genomom, sintetizirani na ribosomima endoplazmatskog retikuluma i transportirani njegovim kanalima do mjesta okupljanja. Tako su mitohondriji rezultat zajedničkih napora dva genoma i dva stroja za transkripciju i translaciju. Neke podjedinice enzima mitohondrijskog respiratornog lanca sastavljene su od različitih polipeptida, od kojih su neki kodirani nuklearnim, a neki mitohondrijskim genomom. Na primjer, ključni enzim oksidativne fosforilacije, citokrom c oksidaza u kvascu, sastoji se od tri podjedinice kodirane i sintetizirane u mitohondrijima, te četiri kodirane u jezgri stanice i sintetizirane u citoplazmi. Ekspresiju većine mitohondrijskih gena kontroliraju specifični geni u jezgri.
Simbiotska teorija o postanku mitohondrija
Hipotezu o podrijetlu mitohondrija i biljnih plastida iz unutarstaničnih bakterija-endosimbiona iznio je R. Altman još 1890. Tijekom stoljeća brzog razvoja biokemije, citologije, genetike i molekularne biologije koja se pojavila prije pola stoljeća, hipoteza je porasla u teoriju koja se temelji na velikoj količini činjeničnog materijala ... Njegova je bit sljedeća: pojavom fotosintetizirajućih bakterija kisik se nakupio u Zemljinoj atmosferi - nusprodukt njihova metabolizma. S povećanjem njegove koncentracije život anaerobnih heterotrofa postao je teži, a neki od njih su, radi dobivanja energije, prešli iz anoksične fermentacije u oksidativnu fosforilaciju. Takvi aerobni heterotrofi mogli bi, s većom učinkovitošću od anaerobnih bakterija, razgraditi organsku tvar nastalu fotosintezom. Neki od slobodnih aeroba uhvaćeni su anaerobima, ali nisu "probavljeni", već su sačuvani kao energetske stanice, mitohondriji. Mitohondrije ne treba promatrati kao robove koji su zarobljeni radi opskrbe stanica molekulama ATP -a koje ne mogu disati. Umjesto toga, oni su "stvorenja" koja su, još u proterozoiku, pronašla najbolja skloništa za sebe i svoje potomke, gdje možete uložiti najmanje truda, a da ne riskirate da vas pojedu.
Brojne činjenice govore u prilog simbiotičkoj teoriji:
Veličine i oblici mitohondrija i slobodnih živućih aerobnih bakterija su isti; obje sadrže kružne molekule DNA koje nisu povezane s histonima (za razliku od linearne nuklearne DNA);
U smislu nukleotidnih sekvenci, ribosomske i transportne RNA mitohondrija razlikuju se od nuklearnih, dok pokazuju iznenađujuće sličnosti sa sličnim molekulama nekih aerobnih gram-negativnih eubakterija;
Mitohondrijske RNA polimeraze, iako kodirane u staničnoj jezgri, inhibirane su rifampicinom, poput bakterijskih, a eukariotske RNA polimeraze su neosjetljive na ovaj antibiotik;
Sintezu proteina u mitohondrijima i bakterijama potiskuju isti antibiotici koji ne utječu na ribosome eukariota;
Sastav lipida unutarnje membrane mitohondrija i bakterijske plazmaleme sličan je, ali se vrlo razlikuje od sastava vanjske membrane mitohondrija, koja je homologna drugim membranama eukariotskih stanica;
Cristae, nastale unutarnjom mitohondrijskom membranom, evolucijski su analozi mezosomalnih membrana mnogih prokariota;
Do sada su organizmi preživjeli koji oponašaju srednje oblike na putu stvaranja mitohondrija iz bakterija (primitivna ameba Pelomyxa nema mitohondrije, ali uvijek sadrži endosimbiotske bakterije).
Postoji ideja da su različita carstva eukariota imala različite pretke, a endosimbioza bakterija je nastala u različitim fazama evolucije živih organizama. O tome svjedoče i razlike u strukturi mitohondrijskih genoma protozoa, gljiva, biljaka i viših životinja. No u svim slučajevima glavni dio gena iz promitohondrija ušao je u jezgru, vjerojatno uz pomoć pokretnih genetskih elemenata. Kada se dio genoma jednog od simbionta uključi u genom drugog, integracija simbionta postaje nepovratna. Novi genom može stvoriti metaboličke putove koji vode do stvaranja korisnih proizvoda koje ne može sintetizirati niti jedan od partnera pojedinačno. Dakle, sinteza steroidnih hormona stanicama kore nadbubrežne žlijezde složen je lanac reakcija, od kojih se neke javljaju u mitohondrijima, a neke u endoplazmatskom retikulumu. Hvatanjem gena promitohondrija jezgra je mogla pouzdano kontrolirati funkcije simbionta. Jezgra kodira sve proteine i sintezu lipida vanjske membrane mitohondrija, većinu proteina matriksa i unutarnje membrane organela. Ono što je najvažnije, jezgra kodira enzime replikacije, transkripcije i translacije mtDNA, kontrolirajući tako rast i reprodukciju mitohondrija. Stopa rasta simbiotskih partnera trebala bi biti približno ista. Ako domaćin raste brže, onda će se sa svakom generacijom broj simbionta po pojedincu smanjivati, a na kraju će se pojaviti potomci bez mitohondrija. Znamo da svaka stanica organizma koja se spolno razmnožava sadrži mnogo mitohondrija koji repliciraju njihovu DNK u razmaku između podjela domaćina. Time se osigurava da će svaka od stanica kćeri primiti barem jednu kopiju mitohondrijskog genoma.
Uloga jezgre stanice u biogenezi mitohondrija
Određena vrsta mutiranog kvasca ima opsežnu deleciju u mitohondrijskoj DNA, što dovodi do potpunog prestanka sinteze proteina u mitohondrijima; zbog toga ti organeli ne mogu obavljati svoju funkciju. Budući da, kad rastu na mediju s niskim udjelom glukoze, takvi mutanti tvore male kolonije, zovu se citoplazmatski mutantamitanka.
Iako mali mutanti nemaju sintezu mitohondrijskih proteina i stoga ne tvore normalne mitohondrije, takvi mutanti ipak sadrže promitohondrije, koji su u određenoj mjeri slični običnim mitohondrijima, imaju normalnu vanjsku membranu i unutarnju membranu sa slabo razvijenim kristama. U promitohondrijama ima mnogo enzima kodiranih nuklearnim genima i sintetiziranih na ribosomima citoplazme, uključujući DNA i RNA polimeraze, sve enzime ciklusa limunske kiseline i mnoge proteine koji čine unutarnju membranu. To jasno pokazuje dominantnu ulogu nuklearnog genoma u biogenezi mitohondrija.
Zanimljivo je napomenuti da, iako izgubljeni fragmenti DNK čine 20 do više od 99,9% mitohondrijskog genoma, ukupna količina mitohondrijske DNA u sitnim mutantima uvijek ostaje na istoj razini kao u divljem tipu. To je posljedica još uvijek slabo proučenog procesa amplifikacije DNA, zbog čega nastaje molekula DNA, koja se sastoji od tandem ponavljanja iste regije i veličine jednake normalnoj molekuli. Na primjer, mitohondrijska DNA malenog mutanta, koja zadržava 50% nukleotidne sekvence DNK divljeg tipa, sastojat će se od dva ponavljanja, dok se molekula koja zadržava samo 0,1% genom divljeg tipa bit će izgrađen od 1000 kopija preostalog fragmenta. Tako se sitni mutanti mogu koristiti za dobivanje u velikom broju određenih dijelova mitohondrijske DNA, koje bi, moglo bi se reći, klonirala sama priroda.
Premda biogenezu organela kontroliraju uglavnom nuklearni geni, same organele, sudeći prema nekim podacima, imaju neku vrstu regulatornog utjecaja na načelo povratne sprege; u svakom slučaju, to je slučaj s mitohondrijima. Ako blokirate sintezu proteina u mitohondrijima netaknutih stanica, tada se enzimi uključeni u mitohondrijsku sintezu DNK, RNK i proteina počinju stvarati višak u citoplazmi, kao da stanica pokušava nadvladati učinak sredstva koje blokira. No, iako postoji sumnja u postojanje neke vrste signala iz mitohondrija, njegova priroda još uvijek nije poznata.
Iz brojnih razloga, mehanizmi biogeneze mitohondrija sada se u većini slučajeva proučavaju u kulturama Saccharomyces carlsbergensis(pivski kvasac i S. cerevisiae(pekarski kvasac). Prvo, kada rastu na glukozi, ti kvasci pokazuju jedinstvenu sposobnost postojanja samo putem glikolize, odnosno bez mitohondrijske funkcije. To omogućuje proučavanje mutacija u mitohondrijskoj i nuklearnoj DNA koje sprječavaju razvoj ovih organela. Takve su mutacije smrtonosne u gotovo svim drugim organizmima. Drugo, kvasac - jednostavni jednostanični eukarioti - lako se uzgaja i biokemijski testira. Konačno, kvasac se može razmnožavati i u haploidnoj i u diploidnoj fazi, obično aseksualnim procesom pupanja (asimetrična mitoza). No, kvasac također ima spolni proces: s vremena na vrijeme se dvije haploidne stanice spoje, tvoreći diploidnu zigotu, koja se tada ili dijeli mitozom ili prolazi kroz mejozu i opet daje haploidne stanice. Kontrolirajući izmjenu aseksualne i spolne reprodukcije tijekom pokusa, možete naučiti mnogo o genima odgovornim za funkciju mitohondrija. Pomoću ovih metoda moguće je, osobito, saznati jesu li takvi geni lokalizirani u nuklearnoj ili mitohondrijskoj DNK, budući da se mutacije mitohondrijskih gena ne nasljeđuju prema Mendelovim zakonima koji uređuju nasljeđivanje nuklearnih gena.
Mitohondrijski transportni sustavi
Većina proteina sadržanih u mitohondrijima i kloroplastima uvozi se u te organele iz citosola. To postavlja dva pitanja: kako stanica usmjerava proteine na odgovarajuću organelu i kako ti proteini u nju ulaze?
Djelomičan odgovor dobiven je proučavanjem transporta male podjedinice (S) enzima u stromu kloroplasta ribuloza-1,5-bisfosfat-karboksilijen. Ako se mRNA izolira iz citoplazme jednostanične alge Chlamydomonas ili iz lišća graška, unesenog kao matrica u sustav za sintezu proteina in vitro, tada će jedan od mnogih nastalih proteina biti vezan specifičnim anti-S-antitijelom. S-protein, sintetiziran in vitro, naziva se pro-S, budući da je veći od običnog S-proteina za oko 50 aminokiselinskih ostataka. Kada se pro-S protein inkubira s netaknutim kloroplastima, on prodire u organele i tamo se peptidazom pretvara u S-protein. Tada se S-protein veže za veliku podjedinicu ribuloza-1,5-bisfosfat karboksilaze, sintetiziranu na ribosomima kloroplasta, i s njom stvara aktivni enzim u stromi kloroplasta.
Mehanizam prijenosa S-proteina nije poznat. Vjeruje se da se pro-S veže na receptorski protein koji se nalazi na vanjskoj membrani kloroplasta ili na mjestu kontakta vanjske i unutarnje membrane, a zatim se transportira u stromu transmembranskim kanalima kao rezultat energije- intenzivan proces.
Proteini se na sličan način transportiraju u mitohondrije. Ako se pročišćene mitohondrije kvasca inkubiraju sa staničnim ekstraktom koji sadrži novo sintetizirane radioaktivne proteine kvasca, može se primijetiti da su mitohondrijski proteini kodirani nuklearnim genomom odvojeni od ne-mitohondrijskih proteina citoplazme i selektivno ugrađeni u mitohondrije, baš kao i oni u netaknutoj ćeliji. U tom slučaju, proteini vanjske i unutarnje membrane, matriksa i intermembranskog prostora nalaze svoj put do odgovarajućeg odjeljka za mitohondrije.
Mnogi od novosintetiziranih proteina namijenjenih unutarnjoj membrani, matrici i međumembranskom prostoru imaju vodeći peptid na svom N-kraju, koji se cijepa specifičnom proteazom u matrici tijekom transporta. Prijenos proteina u ta tri mitohondrijska odjeljka zahtijeva energiju elektrokemijskog gradijenta protona stvorenog preko unutarnje membrane. Mehanizam prijenosa proteina za vanjsku membranu je drugačiji: u ovom slučaju nije potrebna niti potrošnja energije niti proteolitičko cijepanje dužeg proteina prekursora. Ova i druga zapažanja sugeriraju da se sve četiri skupine mitohondrijskih proteina transportiraju u organelu pomoću sljedećeg mehanizma: pretpostavlja se da su svi proteini, osim onih namijenjenih vanjskoj membrani, uključeni u unutarnju membranu kao rezultat procesa koja zahtijeva energiju i javlja se na dodirnim mjestima vanjske i unutarnje membrane. Očigledno, nakon ovog početnog ugradnje proteina u membranu, on prolazi kroz proteolitičko cijepanje, što dovodi do promjene njegove konformacije; ovisno o tome kako se konformacija mijenja, protein je ili fiksiran u membrani ili "gurnut" u matriks ili u međumembranski prostor.
Prijenos proteina kroz membrane mitohondrija i kloroplasta u načelu je analogan njihovom prijenosu kroz membrane endoplazmatskog retikuluma. Međutim, ovdje postoji nekoliko važnih razlika. Prvo, kada se transportira u matriks ili stromu, protein prolazi kroz vanjsku i unutarnju membranu organele, dok prilikom transporta u lumen endoplazmatskog retikuluma molekule prolaze samo kroz jednu membranu. Osim toga, prijenos proteina u retikulum provodi se pomoću mehanizma smjerno uklanjanje(vektorski iscjedak) - počinje kada protein još nije potpuno napustio ribosom (ko-translacijski uvoz), a prijenos u mitohondrije i kloroplaste događa se nakon što je sinteza proteinske molekule potpuno dovršena (posttranslacijski uvoz).
Unatoč tim razlikama, u oba slučaja stanica sintetizira proteine prekursore koji sadrže signalnu sekvencu koja određuje na koju je membranu dani protein usmjeren. Očigledno, u mnogim slučajevima ovaj se slijed cijepa iz molekule prekursora nakon završetka transportnog procesa. Međutim, neki se proteini odmah sintetiziraju u konačnom obliku. Vjeruje se da je u takvim slučajevima signalna sekvenca sadržana u polipeptidnom lancu konačnog proteina. Signalne sekvence još su slabo razumljive, ali vjerojatno mora postojati nekoliko vrsta takvih sekvenci, od kojih svaka određuje prijenos proteinske molekule u određenu regiju stanice. Na primjer, u biljnoj stanici neki se proteini, čija sinteza započinje u citosolu, zatim transportiraju u mitohondrije, drugi u kloroplaste, treći u peroksisom, a treći u endoplazmatski retikulum. Složeni procesi koji vode ispravnoj unutarstaničnoj raspodjeli proteina tek se shvaćaju.
Osim nukleinskih kiselina i proteina, za izgradnju novih mitohondrija potrebni su i lipidi. Za razliku od kloroplasta, mitohondriji većinu lipida primaju izvana. U životinjskim stanicama fosfolipidi sintetizirani u endoplazmatskom retikulumu transportiraju se do vanjske membrane mitohondrija uz pomoć posebnih proteina, a zatim se ugrađuju u unutarnju membranu; vjeruje se da se to događa na mjestu dodira dviju membrana. Glavna reakcija biosinteze lipida, koju kataliziraju sami mitohondriji, je pretvaranje fosfatidne kiseline u fosfolipidni kardiolipin, koji se nalazi uglavnom u unutarnjoj membrani mitohondrija i čini oko 20% svih njegovih lipida.
Veličina i oblik mitohondrijskih genoma
Do danas je pročitano više od 100 različitih mitohondrijskih genoma. Skup i broj njihovih gena u mitohondrijskoj DNA, za koje je nukleotidna sekvenca u potpunosti određena, uvelike se razlikuju kod različitih vrsta životinja, biljaka, gljiva i protozoa. Najveći broj gena pronađenih u mitohondrijskom genomu flagelatnih protozoa Rectinomo-nas americana- 97 gena, uključujući sve gene koji kodiraju proteine koji se nalaze u mtDNA drugih organizama. U većine viših životinja mitohondrijski genom sadrži 37 gena: 13 za proteine respiratornog lanca, 22 za tRNA i dva za rRNA (za veliku podjedinicu ribosoma 16S rRNA i za malu 12S rRNA). U biljkama i protozoama, za razliku od životinja i većine gljiva, neki proteini koji čine ribosome ovih organela kodirani su u mitohondrijskom genomu. Ključni enzimi sinteze predložaka polinukleotida, poput DNK polimeraze (replicirajuća mitohondrijska DNA) i RNA polimeraze (koja prepisuje mitohondrijski genom), kodirani su u jezgri i sintetizirani na citoplazmatskim ribosomima. Ova činjenica ukazuje na relativnost mitohondrijske autonomije u složenoj hijerarhiji eukariotske stanice.
Genomi mitohondrija različitih vrsta razlikuju se ne samo po skupu gena, redoslijedu njihovog smještaja i ekspresije, već i po veličini i obliku DNK. Ogromna većina danas opisanih mitohondrijskih genoma su kružne supermotane dvolančane molekule DNA. U nekim biljkama, uz kružne oblike, postoje i linearni, a u nekim protozoama, na primjer, cilijate, u mitohondrijima se nalazi samo linearna DNA.
U pravilu svaki mitohondrij sadrži nekoliko kopija svog genoma. Dakle, u stanicama ljudske jetre postoji oko 2 tisuće mitohondrija, a u svakoj od njih ima 10 identičnih genoma. U mišjim fibroblastima postoji 500 mitohondrija koji sadrže dva genoma, a u stanicama kvasca S. cerevisiae- do 22 mitohondrija s četiri genoma.
DIV_ADBLOCK1003 ">
 Slika 2. Shema stvaranja linearnih (A), kružnih (B), lančanih (C) oligomera mtDNA. ori - regija početka replikacije DNK.
Slika 2. Shema stvaranja linearnih (A), kružnih (B), lančanih (C) oligomera mtDNA. ori - regija početka replikacije DNK.
Veličina genoma mitohondrija različitih organizama kreće se od manje od 6 tisuća parova baza u plazmodiju malarije (osim dva gena rRNA, sadrži samo tri gena koji kodiraju proteine) do stotina tisuća parova nukleotida u kopnenim biljkama (na primjer, u Arabidopsis thaliana iz obitelji krstašica 366924 parova baza). Istodobno, 7-8 puta se razlikuju u veličini mtDNA viših biljaka čak i unutar iste obitelji. Duljina mtDNA kralježnjaka neznatno se razlikuje: kod ljudi - 16.569 parova baza, kod svinja - 16350, kod dupina - 16330, kod kandži žabe Xenopus laevis- 17533, u šarana- 16400. Ovi su genomi također slični po lokalizaciji gena, od kojih se većina nalazi s kraja na kraj; u nekim slučajevima čak se i preklapaju, obično za jedan nukleotid, tako da je posljednji nukleotid jednog gena prvi u sljedećem. Za razliku od kralježnjaka, u biljkama, gljivama i protozoama, mtDNA sadrže do 80% nekodirajućih sekvenci. Redoslijed gena u genomima mitohondrija razlikuje se od vrste do vrste.
Visoka koncentracija reaktivnih vrsta kisika u mitohondrijima i slab sustav popravka povećavaju učestalost mutacija mtDNA u usporedbi s nuklearnom za red veličine. Radikali kisika uzrokuju specifične zamjene C®T (deaminacija citozina) i GT®T (oksidativno oštećenje gvanina), uslijed čega je, vjerojatno, mtDNA bogata AT-parovima. Osim toga, sve mtDNA imaju zanimljivo svojstvo - nisu metilirane, za razliku od nuklearne i prokariotske DNA. Poznato je da je metilacija (privremena kemijska modifikacija nukleotidne sekvence bez narušavanja kodirajuće funkcije DNA) jedan od mehanizama programirane inaktivacije gena.
Veličina i struktura molekula DNA u organelama
Struktura | Masa, mln. dalton | Bilješke (uredi) |
||
|
ohhon drya | Životinje | Prstenasta | Svaka vrsta ima sve molekule iste veličine |
|
Viši ras sthenia | Prstenasta | Varira | Sve proučavane vrste imaju kružnu DNK različite veličine, u kojoj ukupni sadržaj genetskih informacija odgovara masi od 300 do 1000 milijuna daltona, ovisno o vrsti |
|
Gljive: Najjednostavniji | Prstenasta Prstenasta Prstenasta Linearno | |||
|
Klor opla stov | Alge | Prstenasta Prstenasta | ||
Viši bilje | Prstenasta | Svaka vrsta ima samo jednu molekulu |
Relativna količina DNK organela u nekim stanicama i tkivima
Organizam | Tkanina ili tip ćelije | Broj mol-l DNA / organela | Broj organa- nell u kavez | Udio DNK organela u svemu Stanična DNA,% |
|
|
ohhon drya | |||||
Ćelijska linija L | |||||
Jaje | |||||
|
Klor opla stov | Vegetativne diploidne stanice | ||||
Kukuruz |
Funkcioniranje mitohondrijskog genoma
Što je posebno u mehanizmima replikacije i transkripcije mitohondrijske DNA sisavaca?
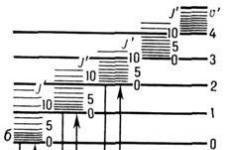
Komplementarne "href =" / text / category / komplementarij / "rel =" bookmark "> komplementarne niti u mtDNA značajno se razlikuju po specifičnoj težini, jer sadrže različite količine" teških "purinskih i" lakih "pirimidinskih nukleotida. - H (teški -teški) i lanac L (lagani-lagani) .Na početku replikacije molekule mtDNA nastaje takozvana D-petlja (iz engleske Displace-ment loop) Ta struktura, vidljiva u elektroničkom mikro -osp, sastoji se od dvolančanih i jednolančanih (uvučeni dio H-lanca) presjeka.kraj sjemena ribonukleotida, što odgovara točki početka sinteze H-lanca (oriH). Sinteza L-lanca započinje tek kad kćer H-lanac dosegne točku ori L. To je zbog činjenice da je regija iniciranja replikacije L-lanca dostupna enzimima za sintezu DNA samo u jednolančanom stanju, i, posljedično, samo u iskrivljenoj dvostrukoj spirali u sintezi H-lanca. Tako se kćeri niti mtDNA sintetiziraju kontinuirano i asinkrono (slika 3).
Slika 3. Shema replikacije mtDNA sisavaca. Prvo se formira D-petlja, zatim se sintetizira kćer H-lanac, zatim započinje sinteza kćeri L-lanca.
Kraj gena 16S rRNA (slika 4). Takvih kratkih prijepisa ima 10 puta više nego dugih. Kao rezultat sazrijevanja (prerade), iz njih nastaju 12S rRNA i 16S rRNA, koje sudjeluju u stvaranju mitohondrijskih ribosoma, kao i fenilalanina i valin tRNA. Preostale tRNA se izrežu iz dugih transkripata i nastaju prevedene mRNA, za čije su 3 "krajeve vezane poliadenilne sekvence. 5" krajevi ovih mRNA nisu prekriveni, što je neuobičajeno za eukariote. Spajanje (spajanje) se ne događa, jer niti jedan od mitohondrijskih gena sisavaca ne sadrži introne.
Slika 4. Transkripcija humane mtDNA koja sadrži 37 gena. Svi se transkripti počinju sintetizirati u regiji ori H. Ribosomske RNA se izrezuju iz transkripata dugog i kratkog H lanca. tRNA i mRNA nastaju kao rezultat obrade iz transkripata oba lanca DNA. Geni tRNA prikazani su svijetlozeleno.
Želite li znati koja još iznenađenja može predstavljati mitohondrijski genom? Fino! Nastavi čitati! ..
Vodeće i 3'-nekodirajuće regije, poput većine nuklearnih mRNA. Brojni geni također sadrže introne. Dakle, u okvirnom genu koji kodira citokrom oksidazu b postoje dva introna. Iz primarnog transkripta RNA autokatalitički (bez sudjelovanja bilo kojeg ili proteini) kopija većine prvog introna je izrezana. Preostala RNA služi kao predložak za stvaranje enzima maturaze, koja je uključena u spajanje. Dio njegove aminokiselinske sekvence kodiran je u preostalim kopijama introna Maturaza ih izrezuje, uništavajući vlastitu mRNA, kopije egzona se spajaju i tvori se mRNA za citokrom oksidazu b (slika 5.) Otkriće ovog fenomena natjeralo nas je da ponovno razmotrimo koncept introna kao „ništa što kodira sekvence ”.
Slika 5. Obrada (sazrijevanje) mRNA citokrom oksidaze b u mitohondrijima kvasca. U prvoj fazi spajanja nastaje mRNA, kojom se sintetizira maturaza, koja je neophodna za drugu fazu spajanja.
Prilikom proučavanja ekspresije mitohondrijskih gena Trypanosoma brucei otkrio iznenađujuće odstupanje od jednog od osnovnih aksioma molekularne biologije koji kaže da se slijed nukleotida u mRNA točno podudara s onim u kodirajućim regijama DNA. Pokazalo se da se mRNA jedne od podjedinica citokrom c oksidaze uređuje, odnosno nakon transkripcije mijenja se njezina primarna struktura - ubacuju se četiri uracila. Kao rezultat toga nastaje nova mRNA, koja služi kao matrica za sintezu dodatne podjedinice enzima, niz aminokiselina u kojem nema nikakve veze sa slijedom. Virus "href =" / text / category / virus / "rel =" bookmark "> virusi, gljivice, Engleski istraživač Burrell usporedio je strukturu jednog od mitohondrijskih gena teleta sa sekvencom aminokiselina u podjedinici citokrom oksidaze koju kodira ovaj gen." idealan je ", odnosno poštuje sljedeće pravilo: „ako dva kodona imaju dva identična nukleotida, a treći nukleotidi pripadaju istoj klasi (purin - A, G ili pirimidin - Y, C), tada kodiraju istu aminokiselinu. ”Postoje dvije iznimke od ovog pravila u univerzalnom kodu: AUA triplet kodira izoleucin, a AUG kodon kodira metionin, dok su u idealnom mitohondrijskom kodu oba ova tripleta do oder metionin; UGG triplet kodira samo triptofan, a UGA triplet kodira stop kodon. U univerzalnom kodu oba odstupanja odnose se na temeljne trenutke sinteze proteina: AUG kodon je inicijator, a UGA stop kodon zaustavlja sintezu polipeptida. Idealni kod nije svojstven svim opisanim mitohondrijima, ali niti jedan od njih nema univerzalni kod. Može se reći da mitohondriji govore različitim jezicima, ali nikada ne govore jezikom jezgre.
Razlike između “univerzalnog” genetskog koda i dva mitohondrijska koda
Codon | Mitohondrijski kod sisavaca | Mitohondrijski kod kvasca | “ Univerzalna” |
Kao što je već spomenuto, u mitohondrijskom genomu kralježnjaka postoje 22 gena tRNA. Kako takav nepotpuni skup služi svih 60 kodona za aminokiseline (u idealnom kodu od 64 trojke postoje četiri stop kodona, u univerzalnom - tri)? Činjenica je da se tijekom sinteze proteina u mitohondrijima interakcije kodon -antikodon pojednostavljuju - dva od tri antikodon nukleotida koriste se za prepoznavanje. Dakle, jedna tRNA prepoznaje sva četiri člana obitelji kodona, koji se razlikuju samo u trećem nukleotidu. Na primjer, leucinska tRNA s GAU antikodonom stoji na ribosomu protiv kodona CUU, CUC, CUA i CUG, osiguravajući ugradnju leucina bez grešaka u polipeptidni lanac. Druga dva leucinska kodona, UUA i UUG, prepoznaju tRNA s antikodonskim AAU. Ukupno osam različitih molekula tRNA prepoznaje osam obitelji od po četiri kodona, a 14 tRNA prepoznaje različite parove kodona, od kojih svaka šifrira jednu aminokiselinu.
Važno je da su enzimi aminoacil tRNA sintetaze odgovorni za vezanje aminokiselina na odgovarajuću tRNA mitohondrija kodirani u jezgri stanice i sintetizirani na ribosomima endoplazmatskog retikuluma. Tako su u kralježnjaka sve proteinske komponente sinteze mitohondrijskog polipeptida kodirane u jezgri. Istodobno, sintezu proteina u mitohondrijima ne potiskuje cikloheksimid, koji blokira rad eukariotskih ribosoma, ali je osjetljiv na antibiotike eritromicin i kloramfenikol, koji inhibiraju sintezu proteina u bakterijama. Ova činjenica služi kao jedan od argumenata u prilog nastanku mitohondrija iz aerobnih bakterija tijekom simbiotskog stvaranja eukariotskih stanica.
Važnost posjedovanja vlastitog genetskog sustava za mitohondrije
Zašto mitohondrije trebaju vlastiti genetski sustav, dok ga drugi organeli, poput peroksisoma i lizosoma, nemaju? Ovo pitanje nije nimalo trivijalno, budući da je održavanje odvojenog genetskog sustava skupo za stanicu, s obzirom na potreban broj dodatnih gena u nuklearnom genomu. Ovdje treba kodirati ribosomske proteine, aminoacil-tRNA sintetaze, DNA i RNA polimeraze, enzime za obradu i modifikaciju RNA itd. Većina proučavanih proteina iz mitohondrija razlikuje se u aminokiselinskom slijedu od njihovih analoga iz drugih dijelova stanice, a tamo razlog je vjerovati da u tim organima postoji vrlo malo proteina koji bi se mogli naći drugdje. To znači da samo za održavanje genetskog sustava mitohondrija nuklearni genom mora imati nekoliko desetaka dodatnih gena. Razlozi za ovaj "otpad" nisu jasni, a nada da će se pronaći trag u nukleotidnoj sekvenci mitohondrijske DNA nije bila opravdana. Teško je zamisliti zašto se proteini nastali u mitohondrijima moraju sintetizirati tamo, a ne u citosolu.
Obično se postojanje genetskog sustava u energetskim organelama objašnjava činjenicom da su neki proteini sintetizirani unutar organele previše hidrofobni da bi mogli proći kroz mitohondrijsku membranu izvana. Međutim, proučavanje kompleksa ATP-sintetaze pokazalo je da je takvo objašnjenje nevjerojatno. Iako su pojedinačne proteinske podjedinice ATP sintetaze tijekom evolucije vrlo očuvane, mjesta njihove sinteze se mijenjaju. U kloroplastima se na ribosomima unutar organele stvara nekoliko prilično hidrofilnih proteina, uključujući četiri od pet podjedinica dijela kompleksa F1-ATPaze. Naprotiv, kod gljive Neurospora a u životinjskim stanicama se na citoplazmatskim ribosomima sintetizira vrlo hidrofobna komponenta (podjedinica 9) membranskog dijela ATPaze i tek nakon toga prelazi u organelu. Različitu lokalizaciju gena koji kodiraju podjedinice funkcionalno ekvivalentnih proteina u različitim organizmima teško je objasniti uz pomoć bilo koje hipoteze koja pretpostavlja određene evolucijske prednosti suvremenih genetskih sustava mitohondrija i kloroplasta.
Uzimajući u obzir sve navedeno, ostaje samo pretpostaviti da genetski sustav mitohondrija predstavlja evolucijsku slijepu ulicu. U okviru endo-simbiotske hipoteze, to znači da je proces prijenosa endosimbiotskih gena u nuklearni genom domaćina stao prije nego što je potpuno završen.
Citoplazmatsko nasljeđivanje
Posljedice prijenosa citoplazmatskih gena za neke životinje, uključujući ljude, ozbiljnije su nego za kvasac. Dvije haploidne stanice kvasca koje se spajaju iste su veličine i doprinose istoj količini mitohondrijske DNA rezultirajućoj zigoti. Dakle, u kvascu je mitohondrijski genom naslijeđen od oba roditelja, koji daju jednak doprinos genskom fondu potomaka (iako je nakon nekoliko generacija odvojiti potomci će često sadržavati mitohondrije samo jednog od roditeljskih tipova). Nasuprot tome, kod viših životinja jajna stanica unosi više citoplazme u zigotu od spermija, a kod nekih životinja spermiji uopće ne mogu dodati citoplazmu. Stoga se može pomisliti da će se kod viših životinja mitohondrijski genom prenijeti samo od jednog roditelja (naime majčinski linije); i doista, to je potvrđeno eksperimentima. Pokazalo se, na primjer, da se križanjem štakora dviju laboratorijskih linija s mitohondrijskom DNA neznatno razlikuju u slijedu nukleotida (tipovi A i B) dobiva potomstvo koje sadrži
koja sadrži mitohondrijsku DNA samo majčinskog tipa.
Citoplazmatska nasljednost, za razliku od nuklearne, ne poštuje Mendelove zakone. To je zbog činjenice da u višim životinjama i biljkama gamete različitih spolova sadrže neusporedive količine mitohondrija. Dakle, u jajetu miša ima 90 tisuća mitohondrija, a u spermi - samo četiri. Očito, u oplođenom jajašcu mitohondriji su pretežno ili samo od ženke, tj. Nasljeđivanje svih mitohondrijskih gena je majčino. Genetska analiza citoplazmatske nasljednosti je teška zbog nuklearno-citoplazmatskih interakcija. U slučaju citoplazmatske muške sterilnosti, mutirani mitohondrijski genom stupa u interakciju s određenim nuklearnim genima, čiji su recesivni aleli neophodni za razvoj svojstva. Dominantni aleli ovih gena, i u homo- i u heterozigotnim stanjima, vraćaju plodnost biljaka bez obzira na stanje mitohondrijskog genoma.
Željela bih se osvrnuti na mehanizam nasljeđivanja majčinskih gena dajući konkretan primjer. Kako bismo konačno i neopozivo razumjeli mehanizam nemendelskog (citoplazmatskog) nasljeđivanja mitohondrijskih gena, razmotrimo što se događa s takvim genima kada se dvije haploidne stanice spoje i tvore diploidnu zigotu. U slučaju kada jedna stanica kvasca nosi mutaciju koja određuje otpornost sinteze proteina mitohondrija na kloramfenikol, a druga, stanica divljeg tipa, osjetljiva je na ovaj antibiotik: mutirani geni mogu se lako identificirati uzgojem kvasca na mediju s glicerol, koji mogu koristiti samo stanice s netaknutim mitohondrijima; stoga, u prisutnosti kloramfenikola, samo stanice koje nose mutirani gen mogu rasti na takvom mediju. Naša diploidna zigota u početku će imati i mutantne i mitohondrije divljeg tipa. Kao rezultat mitoze, iz zigote će niknuti diploidna stanica kćer, koja će sadržavati samo mali broj mitohondrija. Nakon nekoliko mitotičkih ciklusa, na kraju će jedna od novih stanica primiti sve mitohondrije, mutantne ili divlje. Stoga će svi potomci takve stanice imati genetski identične mitohondrije. Takav slučajni proces, uslijed kojeg nastaje diploidno potomstvo koje sadrži samo jednu vrstu mitohondrija, naziva se mitotičkith gleGrčkath. Kad diploidna stanica sa samo jednom vrstom mitohondrija doživi mejozu, sve četiri kćeri haploidne stanice primaju iste mitohondrijske gene. Ova vrsta nasljeđivanja naziva se nemendeLav preslikati ili citoplazmatska za razliku od mendelskog nasljeđivanja nuklearnih gena. Prijenos gena po citoplazmatskom tipu znači da se ispitivani geni nalaze u mitohondrijima.
Proučavanje mitohondrijskih genoma, njihove evolucije, prema posebnim zakonima populacijske genetike, odnosa između nuklearnog i mitohondrijskog genetskog sustava, potrebno je za razumijevanje složene hijerarhijske organizacije eukariotske stanice i organizma u cjelini.
Određene nasljedne bolesti i starenje ljudi povezane su s određenim mutacijama u mitohondrijskoj DNA ili u nuklearnim genima koji kontroliraju mitohondrijsku funkciju. Akumuliraju se podaci o sudjelovanju defekata mtDNA u karcinogenezi. Stoga mitohondriji mogu biti meta kemoterapije raka. Postoje činjenice o bliskoj interakciji nuklearnog i mitohondrijskog genoma u razvoju niza ljudskih patologija. Više je delecija mtDNA pronađeno u pacijenata s teškom mišićnom slabošću, ataksijom, gluhoćom, mentalnom retardacijom, naslijeđenom na autosomno dominantan način. Utvrđen spolni dimorfizam u kliničkim manifestacijama koronarne bolesti srca, koji je najvjerojatnije posljedica majčinog učinka - citoplazmatske nasljednosti. Razvoj genske terapije daje nadu za ispravljanje nedostataka u genomima mitohondrija u doglednoj budućnosti.
Kao što znate, kako bi se provjerila funkcija jedne od komponenti višekomponentnog sustava, potrebno je ukloniti ovu komponentu, nakon čega slijedi analiza promjena koje su se dogodile. Budući da je tema ovog sažetka pokazatelj uloge majčinog genoma u razvoju potomaka, bilo bi logično saznati o posljedicama kršenja u sastavu mitohondrijskog genoma uzrokovanim različitim čimbenicima. Pokazalo se da je mutacijski proces alat za proučavanje gornje uloge, a posljedice njegova djelovanja koje nas zanima bile su tzv. mitohondrijske bolesti.
Mitohondrijske bolesti primjer su citoplazmatske nasljednosti kod ljudi, odnosno "nasljedstva organela". Ovo pojašnjenje treba učiniti jer sada je dokazano postojanje, barem u nekim organizmima, citoplazmatskih nasljednih odrednica koje nisu povezane s staničnim organelama - citogenima (Vechtomov, 1996).
Mitohondrijske bolesti heterogena su skupina bolesti uzrokovanih genetskim, strukturnim, biokemijskim oštećenjima mitohondrija i oslabljenim disanjem tkiva. Za dijagnosticiranje mitohondrijske bolesti važna je opsežna genealoška, klinička, biokemijska, morfološka i genetska analiza. Glavni biokemijski znak mitohondrijske patologije je razvoj laktacidoze, obično se otkrije hiperlaktatacidemija u kombinaciji s hiperpiruvatacidemijom. Broj različitih varijanti dosegao je 120 oblika. Postoji stabilan porast koncentracije mliječne i piruvične kiseline u cerebrospinalnoj tekućini.
Mitohondrijske bolesti (MB) predstavljaju značajan problem za suvremenu medicinu. Prema metodama nasljednog prijenosa, među MB -ima razlikuju se bolesti koje se monogeno nasljeđuju prema Mendelskom tipu, u kojima je zbog mutacije nuklearnih gena poremećena ili struktura i funkcioniranje proteina mitohondrija, ili ekspresija promjene mitohondrijske DNA, kao i bolesti uzrokovane mutacijama mitohondrijskih gena, koje se uglavnom prenose na potomstvo po majčinoj liniji.
Podaci morfoloških studija koji ukazuju na grubu patologiju mitohondrija: abnormalna proliferacija mitohondrija, polimorfizam mitohondrija s poremećajima u obliku i veličini, neorganiziranost kristala, nakupine abnormalnih mitohondrija ispod sarkoleme, parakristalne inkluzije u mitohondrijama, prisutnost interfibrilarnih vakuola
Oblici mitohondrijskih bolesti
1 ... Mitohondrijske bolesti uzrokovane mutacijama mitohondrijske DNA
1.1 Bolesti uzrokovane delecijom mitohondrijske DNA
1.1.1. Kearns-Sayreov sindrom
Bolest se očituje u dobi od 4-18 godina, progresivna vanjska oftalmoplegija, pigmentozni retinitis, ataksija, namjerni tremor, atrioventrikularni srčani blok, povećana razina proteina u cerebrospinalnoj tekućini više od 1 g / l, "poderana" crvena vlakna u biopsijama skeletnih mišića
1.1.2 Pearsonov sindrom
Prvijenac bolesti od rođenja ili u prvim mjesecima života, ponekad je moguće razviti encefalomiopatije, ataksiju, demenciju, progresivnu vanjsku oftalmoplegiju, hipoplastičnu anemiju, oslabljenu egzokrinu funkciju gušterače, progresivni tijek
2 Bolesti uzrokovane točkastim mutacijama mitohondrijske DNA
Majčinski tip nasljeđivanja, akutno ili subakutno smanjenje vidne oštrine na jednom ili oba oka, kombinacija s neurološkim i osteoartikularnim poremećajima, retinom mikroangiopatija, progresivni tijek s mogućnošću remisije ili obnove vidne oštrine, početak bolesti u dobi od 20 godina -30 godina
2.2 NAPR sindrom (neuropatija, ataksija, pigmentozni retinitis)
Majčinski tip nasljeđivanja, kombinacija neuropatije, ataksije i pigmentoze retinitisa, odgođen psihomotorni razvoj, demencija, prisutnost "poderanih" crvenih vlakana u biopsijama mišićnog tkiva
2.3. MERRF sindrom (mioklonus-epilepsija, "poderana" crvena vlakna)
Majčinski tip nasljeđivanja, početak bolesti u dobi od 3 do 65 godina, mioklonska epilepsija, ataksija, demencija u kombinaciji s neurosenzornom gluhoćom, atrofija vidnog živca i oslabljena duboka osjetljivost, kupaonski epileptički kompleksi laktatne acidoze, "poderana" crvena vlakna u biopsije skeletnih mišića, progresivni tijek
2.4 MELAS sindrom (mitohondrijska encefalomiopatija, laktacidoza, epizode slične moždanom udaru)
Majčinski tip nasljeđivanja, početak bolesti prije 40. godine, netolerancija na vježbe, glavobolje slične migreni s mučninom i povraćanjem, epizode slične moždanom udaru, konvulzije, laktatna acidoza, "poderana" crvena vlakna u biopsijama mišića, progresivni tijek.
3 .Patologija povezana s nedostacima u međugenomskoj komunikaciji
3.1 Sindromi višestruke delecije mitohondrijske DNA
Blefaroptoza, vanjska oftalmoplegija, mišićna slabost, senzorineuralna gluhoća, optička atrofija, progresivni tijek, "poderana" crvena vlakna u biopsijama skeletnih mišića, smanjena aktivnost enzima dišnog lanca.
3.2 Sindrom delecije mitohondrijske DNA
Autosomno recesivno nasljeđivanje
Klinički oblici:
3.2.1.Fatalno infantilno
a) teško oštećenje jetre b) hepatopatija c) hipotenzija mišića
Debi u neonatalnom razdoblju
3.2.2.Urođena miopatija
Teška mišićna slabost, generalizirana hipotenzija, kardiomiopatija i napadaji, oštećenje bubrega, glukozurija, aminoacidopatija, fosfaturija
3.2.3.Infantilna miopatija
javlja se u prve 2 godine života, progresivna mišićna slabost, atrofija proksimalnih mišićnih skupina i gubitak tetivnih refleksa, brzo progresivni tijek, smrt u prve 3 godine života.
4 .Mitohondrijske bolesti uzrokovane nuklearnim DNK mutacijama
4.1 Bolesti povezane s defektima u respiratornom lancu
4.1.1 Nedostatak kompleksa 1 (NADH: CoQ reduktaza)
Početak bolesti prije 15 godina, sindrom miopatije, usporen psihomotorni razvoj, poremećaj kardiovaskularnog sustava, napadaji otporni na terapiju, više neuroloških poremećaja, progresivni tijek
4.1.2. Nedostatak kompleksa 2 (sukcinat-CoQ reduktaza)
Karakteriziran je sindromom encefalomiopatije, progresivnim tijekom, podputovima, moguć je razvoj ptoze
4.1.3. Nedostatak kompleksa 3 (CoQ-citokrom C-oksidoreduktaza)
Poremećaji više sustava, oštećenja različitih organa i sustava, s zahvaćanjem središnjeg i perifernog živčanog sustava, endokrinog sustava, bubrega, progresivni tijek
4.1.4. Složeni (citokrom C-oksidaza) nedostatak
4.1.4.1 Fatalna infantilna kongenitalna laktacidoza
Mitohondrijska miopatija s zatajenjem bubrega ili kardiomiopatijom, debi u neonatalnoj dobi, teški respiratorni poremećaji, difuzna mišićna hipotenzija, progresivni tijek, smrt u prvoj godini života.
4.1.4.2.Dobroćudna dječja slabost mišića
Atrofija, uz odgovarajuće i pravodobno liječenje, moguća je brza stabilizacija procesa i oporavak do 1-3 godine života
5 .Menkesov sindrom (trihopoliodistrofija)
Naglo kašnjenje u psihomotornom razvoju, zaostajanje u rastu, oslabljen rast i distrofične promjene u kosi,
6 ... Mitohondrijske encefalomiopatije
6.1.Leigh -ov sindrom(subakutna neurotična encefalomijelopatija)
Pojavljuje se nakon 6 mjeseci života, hipotonija mišića, ataksija, nistagmus, piramidalni simptomi, oftalmoplegija, atrofija vidnih živaca, često povezana s kardiomiopatijom i blagom metaboličkom acidozom
6.2.Alpersov sindrom(progresivna sklerozirajuća polidistrofija)
Degeneracija sive tvari mozga u kombinaciji s cirozom jetre, nedostatkom kompleksa 5 (ATP sintetaza), odgođenim psihomotornim razvojem, ataksijom, demencijom, slabošću mišića, progresivnim tijekom bolesti, nepovoljnom prognozom
6.3 Nedostatak koenzima-Q
Metaboličke krize, mišićna slabost i umor, oftalmoplegija, gluhoća, smanjenje vida, epizode slične moždanom udaru, ataksija, epilepsija mioklonusa, oštećenje bubrega: glukozurija, aminoacidopatija, fosfaturija, endokrini poremećaji, progresivni tijek, smanjena aktivnost respiratornih enzima
7 .Bolesti povezane s poremećenim metabolizmom mliječne i piruvične kiseline
7.1. Nedostatak piruvat karboksilaze Autosomno recesivni način nasljeđivanja, početak bolesti u neonatalnom razdoblju, kompleks simptoma "mlitavog djeteta", napadaji otporni na terapiju, visoke koncentracije ketonskih tijela u krvi, hiperamonemija, hiperlizinemija, smanjena aktivnost piruvat karboksilaze u skeletnim mišićima
7.2 Nedostatak piruvat dehidrogenaze
Manifestacija u neonatalnom razdoblju, kraniofacijalna dismorfija, napadaji otporni na terapiju, otežano disanje i sisanje, kompleks simptoma "mlitavo dijete", disginezija mozga, teška acidoza s visokim udjelom laktata i piruvata
7.3 Smanjena aktivnost piruvat dehidrogenaze
Manifestacija u prvoj godini života, mikrocefalija, odgođen psihomotorni razvoj, ataksija, mišićna distonija, koreoatetoza, laktacidoza s visokim udjelom piruvata
7.4 Nedostatak dihidrolipoiltransacetilaze
Autosomno recesivni tip nasljeđivanja, početak bolesti u neonatalnom razdoblju, mikrocefalija, odgođen psihomotorni razvoj, hipotenzija mišića praćena povećanjem mišićnog tonusa, atrofija diska vidnog živca, laktacidoza, smanjena aktivnost dihidrolipoiltrans-acetilaze
7.5 Nedostatak dihidrolipoil dehidrogenaze
Autosomno recesivni tip nasljeđivanja, početak bolesti u prvoj godini života, kompleks simptoma "mlitavog djeteta", dismetaboličke krize s povraćanjem i proljevom, odgođen psihomotorni razvoj, atrofija diska optičkog živca, laktatna acidoza, povišen alanin u serumu, α-ketoglutarat, α-keto kiseline razgranatog lanca, smanjena aktivnost dihidrolipoil dehidrogenaze
8 Bolesti uzrokovane nedostacima beta-oksidacije masnih kiselina
8.1 Nedostatak dugolančane acetil-CoA dehidrogenaze
Autosomno recesivni način nasljeđivanja, početak bolesti u prvim mjesecima života, metaboličke krize s povraćanjem i proljevom, simptomi "mlitavog djeteta", hipoglikemija, dikarboksilna kiselina, smanjena aktivnost acetil-CoA dehidrogenaze masnih kiselina s dugotrajnom ugljikov lanac
8.2 Nedostatak srednje ugljične acetil CoA dehidrogenaze
Autosomno recesivni način nasljeđivanja, početak bolesti u neonatalnom razdoblju ili prvim mjesecima života, metaboličke krize s povraćanjem i proljevom,
mišićna slabost i hipotenzija, često se razvija sindrom iznenadne smrti, hipoglikemija, dikarboksilna acidurija, smanjena acetil-CoA dehidrogenaza masnih kiselina lanca srednjeg ugljika
8.3. Nedostatak acetil-CoA dehidrogenaze kratkih ugljikovih lanaca masnih kiselina
Autosomno recesivno nasljeđivanje, različite dobi od početka bolesti, smanjena tolerancija na vježbanje, metaboličke krize s povraćanjem i proljevom, slabost mišića i hipotenzija, povećano izlučivanje metil jantarne kiseline urinom, acetil-CoA dehidrogenaza masnih kiselina kratkog ugljikovog lanca
8.4 Višestruki nedostatak acetil-CoA dehidrogenaze masnih kiselina
Neonatalni oblik: kraniofacijalna dismorfija, moždana disginezija, teška hipoglikemija i acidoza, maligni tijek, smanjena aktivnost svih acetil-CoA dehidrogenaza masnih kiselina,
Infantilni oblik: usporeni kompleks simptoma djeteta, kardiomiopatija, metaboličke krize, hipoglikemija i acidoza
8.5 Smanjena aktivnost svih acetil-CoA dehidrogenaza masnih kiselina
Kasni debi obrazac: periodične epizode mišićne slabosti, metaboličke krize, hipoglikemija i acidoza manje su izražene, inteligencija je očuvana,
9 .Krebovi ciklusa fermentopatija
9.1 Nedostatak fumaraze
Autosomno recesivni način nasljeđivanja, početak bolesti u neonatalnom ili neonatalnom razdoblju, mikrocefalija, opća mišićna slabost i hipotenzija, epizode letargije, brzo progresivna en-cefalopatija, loša prognoza
9.2 Nedostatak sukcinat dehidrogenaze
Rijetka bolest koju karakterizira progresivna encefalomiopatija
9.3 Nedostatak alfa-ketoglutarat dehidrogenaze
Autosomno recesivni način nasljeđivanja, neonatalni početak bolesti, mikrocefalija, kompleks simptoma "mlitavog djeteta", epizode letargije, laktatna acidoza, brzo progresivni tijek, smanjenje sadržaja enzima Krebsovog ciklusa u tkivima
9.4.Sindromi nedostatka karnitina i enzimi njegovog metabolizma
Nedostatak karnitin palmitoiltransferaze-1, autosomno recesivni način nasljeđivanja, rani početak bolesti, epizode nekinetonemijske hipoglikemijske kome, hepatomegalija, hipertrigliceridemija i umjerena hiperamonijemija, smanjena aktivnost karnitin-palmitoiltransferaze-1 u jetri karnitin-palmitoiltransferaze-1
9.5 Nedostatak karnitin acilkarnitin translokaze
Rani početak bolesti, kardiovaskularni i respiratorni poremećaji, kompleks simptoma "mlitavog djeteta", epizode letargije i kome, povećanje koncentracije estera karnitina i dugačak lanac ugljika na pozadini smanjenja slobodnog karnitina u krvnog seruma, smanjenje aktivnosti karnitin-acilkarnitin-translokaze
9.6 Nedostatak karnitin palmitoiltransferaze-2
Autosomno recesivno nasljeđivanje, slabost mišića, mijalgija, mioglobinurija, smanjena aktivnost karnitin palmitoiltransferaze-2 u skeletnim mišićima
Autosomno recesivni način nasljeđivanja, kompleks miopatskih simptoma, epizode letargije i letargije, kardiomiopatija, epizode hipoglikemije, smanjene razine karnitina u serumu i povećano izlučivanje karnitina mokraćom.
Nakon analize tako 'strašnog' popisa patologija povezanih s određenim promjenama u funkcioniranju mitohondrijskog (i ne samo) genoma, postavljaju se određena pitanja. Koji su proizvodi mitohondrijskih gena i u kojim super-mega-vitalnim staničnim procesima sudjeluju?
Kako se pokazalo, neke od gore navedenih patologija mogu nastati sintezom 7 podjedinica kompleksa NADH dehidrogenaze, 2 podjedinice ATP sintetaze, 3 podjedinice citokrom c oksidaze i 1 podjedinice ubikinol citokrom c reduktaze (citokrom b), koje su genski produkti mitohondrija. Na temelju toga može se zaključiti da za ove proteine postoji ključna uloga u procesima staničnog disanja, oksidacije masnih kiselina i sintezi ATP -a, prijenosu elektrona u transportnom sustavu elektrona unutarnje MT membrane, funkcioniranju antioksidansa sustav itd.
Sudeći prema najnovijim podacima o mehanizmima apoptoze, mnogi su znanstvenici došli do zaključka da postoji centar za kontrolu apoptoze upravo ...
Uloga mitohondrijskih proteina također se pokazala s antibioticima koji blokiraju sintezu MT. Ako se ljudske stanice u kulturi tkiva liječe antibiotikom, poput tetraciklina ili kloramfenikola, tada će nakon jedne ili dvije podjele njihov rast prestati. To je posljedica inhibicije sinteze proteina mitohondrija, što dovodi do pojave defektnih mitohondrija i, kao posljedica, do nedovoljnog stvaranja ATP -a. Zašto se onda antibiotici mogu koristiti za liječenje bakterijskih infekcija? Na ovo pitanje postoji nekoliko odgovora:
1. Neki antibiotici (poput eritromicina) ne prolaze kroz unutarnju membranu mitohondrija sisavaca.
2. Većina stanica u našem tijelu ne dijeli se niti se dijeli vrlo sporo, pa se zamjena postojećih mitohondrija novim događa jednako sporo (u mnogim tkivima polovica mitohondrija zamjenjuje se za otprilike pet dana ili čak i dulje). Tako će se broj normalnih mitohondrija smanjiti na kritičnu razinu samo ako se blokada sinteze mitohondrijskih proteina održava mnogo dana.
3. Određeni uvjeti unutar tkiva sprječavaju prodiranje određenih lijekova u mitohondrije najosjetljivijih stanica. Na primjer, visoka koncentracija Ca2 + u koštanoj srži dovodi do stvaranja kompleksa Ca2 + tetraciklina, koji ne može prodrijeti u brzo diobene (pa stoga i najranjivije) prekursore krvnih stanica.
Ovi čimbenici omogućuju upotrebu nekih lijekova koji inhibiraju sintezu proteina mitohondrija kao antibiotike u liječenju viših životinja. Samo dva od ovih lijekova imaju nuspojave: dugotrajno liječenje velikim dozama kloramfenikola može dovesti do poremećaja hematopoetske funkcije koštane srži (potisnuti stvaranje crvenih krvnih stanica i leukocita), a dugotrajna uporaba tetraciklina može oštetiti crijevni epitel. No u oba slučaja još nije potpuno jasno jesu li ove nuspojave uzrokovane blokadom mitogeneze biogeneze ili nekim drugim razlogom.
Izlaz
Strukturne i funkcionalne značajke mt genoma su sljedeće. Prvo je utvrđeno da se mtDNA prenosi s majke na sve nje
potomci i od njezinih kćeri na sve sljedeće generacije, ali sinovi ne prenose svoju DNK (majčinsko naslijeđe). Majčinski karakter
nasljeđivanje mtDNA vjerojatno je posljedica dvije okolnosti: ili je udio očeve mtDNA tako mali (očinski
više od jedne molekule DNA na 25 tisuća mtDNA majke) koje se ne mogu otkriti postojećim metodama ili je replikacija mitohondrija s očeve strane blokirana nakon oplodnje. Drugo, odsutnost kombinacijske varijabilnosti - mtDNA pripada samo jednom od roditelja, stoga nema rekombinacijskih događaja karakterističnih za nuklearnu DNA u mejozi, a nukleotidna sekvenca mijenja se iz generacije u generaciju samo zbog mutacija. Treće, mtDNA nema introna
(velika vjerojatnost da će slučajna mutacija utjecati na kodirajuće područje DNA), zaštitni histoni i učinkovit sustav popravka DNA - sve to određuje 10 puta veću stopu mutacija nego u nuklearnoj DNA. Četvrto, unutar iste stanice normalna i mutirana mtDNA mogu istodobno koegzistirati - fenomen heteroplazme (prisutnost samo normalne ili samo mutirane mtDNA naziva se homoplazma). Konačno, oba lanca su prepisana i prevedena u mtDNA, a po nizu karakteristika genetski kod mtDNA razlikuje se od univerzalnog (UGA kodira triptofan, AUA kodira metionin, AGA i AGG su zaustavljeni
kodoni).
Ova svojstva i gore spomenute funkcije mt genoma učinile su proučavanje varijabilnosti nukleotidne sekvence mtDNA neprocjenjivim oruđem za liječnike, sudske liječnike, evolucijske biologe,
predstavnici povijesne znanosti u rješavanju svojih specifičnih zadataka.
Od 1988., kada je otkriveno da su mutacije u genima mtDNA u osnovi mitohondrijskih miopatija (JY Holt i sur., 1988.) i Leberova nasljedna optička neuropatija (DC Wallace, 1988.), daljnja sustavna identifikacija mutacija u ljudskom mt genomu dovela je do stvaranja koncepta mitohondrijskih bolesti (MB). Trenutno su patološke mutacije mtDNA otkrivene u svakoj vrsti mitohondrijskih gena.
Bibliografija
1. Skulačev, mitohondriji i kisik, Soros. obrazovan. zhurn.
2. Osnove biokemije: U tri sveska, M.: Mir ,.
3. Nicholes D. G. Bioenergetika, An Introd. do Kemiozma. Th., Akad. Tisak, 1982.
4. Stryer L. Biokemija, 2. izd. San Fransisco, Freeman, 1981.
5. Skulačev bioloških membrana. M., 1989. godine.
6., Čencov retikulum: struktura i neke funkcije // Rezultati znanosti. Opći problemi biologije. 1989. godine
7. Centologija Chentsov. M.: Izdavačka kuća Moskovskog državnog sveučilišta, 1995
8. , Sfera kompetencije mitohondrijskog genoma // Vestn. RAMS, 2001. broj 10, str. 31-43.
9. Holt I. J, Harding A. E., Morgan-Hughes I. A. Brisanje mišićne mitohondrijske DNA mišića u bolesnika s mitohondrijskim miopatijama. Nature 1988, 331: 717-719.
10. i tako dalje. Ljudski genom i geni predispozicije. SPb., 2000
11. , Mitohondrijski genom. Novosibirsk, 1990.
12. // Soros. obrazovan. zhurn. 1999. broj 10. S.11-17.
13. Uloga simbioze u evoluciji stanica. M., 1983. godine.
14. // Soros. obrazovan. zhurn. 1998. broj 8. S.2-7.
15. // Soros. obrazovan. zhurn. 2000. Broj 1. S.32-36.
Kijevsko nacionalno sveučilište. Taras Ševčenko
Odjel za biologiju
sažetak
na temu:
"Uloga majčinog genoma u razvoju potomaka"
sudaratienta IVtečaj
Odjel za biokemiju
Frolova Artem
Kijev 2004
Plan:
Uvod................................................. .............................. 1
Simbiotska teorija podrijetla mitohondrija ... 2
Uloga jezgre stanice u biogenezi mitohondrija ................................... 5
Transportni sustavi mitohondrija .............................................. ...... 7
Veličina i oblik mitohondrijskih genoma .................. 10
Funkcioniranje mitohondrijskog genoma ............... 14
Važnost posjedovanja vlastitog genetskog sustava za mitohondrije ........................................ ... ................................... 19
Citoplazmatsko nasljeđivanje .............................. 20
Povijesno gledano, prva studija ove vrste provedena je pomoću mitohondrijske DNA. Znanstvenici su uzeli uzorak iz starosjedilaca Afrike, Azije, Europe, Amerike i u ovom, u početku malom uzorku, međusobno su uspoređivali mitohondrijsku DNA različitih pojedinaca. Otkrili su da je raznolikost mitohondrijske DNA najveća u Africi. A budući da je poznato da mutacijski događaji mogu promijeniti tip mitohondrijske DNA, a poznato je i kako se može promijeniti, stoga je, dakle, moguće reći koje su vrste ljudi iz kojih su mogle nastati mutacijski. Kod svih ljudi kojima je testiran DNK, kod Afrikanaca je pronađena mnogo veća varijabilnost. Vrste mitohondrijske DNA na drugim kontinentima bile su manje raznolike. To znači da su Afrikanci imali više vremena za akumuliranje ovih promjena. Imali su više vremena za biološku evoluciju, ako se baš u Africi pronađu drevni ostaci DNK koji nisu karakteristični za europske ljudske mutacije.
Može se tvrditi da su genetičari mitohondrijske DNA uspjeli dokazati podrijetlo žena u Africi. Proučavali su i Y kromosome. Pokazalo se da i muškarci dolaze iz Afrike.
Zahvaljujući istraživanju mitohondrijske DNK, moguće je ustanoviti ne samo da je osoba podrijetlom iz Afrike, već i utvrditi vrijeme svog podrijetla. Vrijeme pojavljivanja mitohondrijske pramajke čovječanstva utvrđeno je usporednim istraživanjem mitohondrijske DNA čimpanzi i modernog čovjeka. Poznavajući stopu mutacijske divergencije - 2-4% na milijun godina - moguće je odrediti vrijeme razdvajanja dviju grana, čimpanzi i modernog čovjeka. To se dogodilo prije otprilike 5-7 milijuna godina. U tom se slučaju stopa mutacijske divergencije smatra konstantnom.
Mitohondrijsko predvečerje
Kad ljudi govore o mitohondrijskoj Evi, ne misle na pojedinca. Oni govore o evoluciji čitave populacije pojedinaca sa sličnim osobinama. Vjeruje se da je mitohondrijska Eva živjela u razdoblju naglog pada broja naših predaka, na oko deset tisuća jedinki.
Podrijetlo rasa
Proučavajući mitohondrijsku DNK različitih populacija, genetičari su sugerirali da je čak i prije napuštanja Afrike populacija predaka podijeljena u tri skupine, dajući tako tri moderne rase - afričku, bijelu i mongoloidnu. Vjeruje se da se to dogodilo prije otprilike 60 - 70 tisuća godina.
Usporedba mitohondrijske DNA nestandardnih i modernih ljudi
Dodatne informacije o podrijetlu ljudi dobivene su usporedbom genetskih tekstova mitohondrijske DNA neandertalca i modernog čovjeka. Znanstvenici su mogli čitati genetske tekstove mitohondrijske DNK koštanih ostataka dvaju neandertalaca. Kosti prvog neandertalca pronađene su u pećini Feldhover u Njemačkoj. Nešto kasnije pročitan je genetski tekst mitohondrijske DNK neandertalskog djeteta koji je pronađen na Sjevernom Kavkazu u pećini Mezhmayskaya. Uspoređujući mitohondrijsku DNA modernog čovjeka i neandertalca, pronađene su vrlo velike razlike. Uzmemo li dio DNK, tada se od 370 nukleotida razlikuje 27. A ako usporedimo genetske tekstove modernog čovjeka, njegovu mitohondrijsku DNA, tada se razlikuje samo osam nukleotida. Vjeruje se da su neandertalac i moderni čovjek potpuno odvojene grane, evolucija svake od njih odvijala se neovisno jedna o drugoj.
Proučavajući razliku u genetskim tekstovima mitohondrijske DNK neandertalca i modernog čovjeka, utvrđen je datum razdvajanja ove dvije grane. To se dogodilo prije otprilike 500 tisuća godina, a prije otprilike 300 tisuća godina dogodilo se njihovo konačno razdvajanje. Vjeruje se da su se neandertalci naselili u Europi i Aziji, a istjerao ih je čovjek modernog tipa koji je 200 tisuća godina kasnije napustio Afriku. I konačno, prije otprilike 28 - 35 tisuća godina, neandertalci su izumrli. Zašto se to uopće dogodilo, još nije jasno. Možda nisu mogli podnijeti konkurenciju s osobom modernog tipa, ili su za to postojali drugi razlozi.
© G.M. Dymshits
Mitohondrijski genom iznenađuje
G.M. Gluposti
Grigorij Moisejevič Dymshits, Doktor bioloških znanosti, profesor Odjela za molekularnu biologiju Novosibirskog državnog sveučilišta, voditelj laboratorija za strukturu genoma Instituta za citologiju i genetiku Sibirskog ogranka Ruske akademije znanosti. Koautor i urednik četiri školska udžbenika iz opće biologije.Od otkrića molekula DNA u mitohondrijima prošlo je četvrt stoljeća prije nego što su se zainteresirali ne samo za molekularne biologe i citologe, već i za genetičare, evolucioniste, kao i za paleontologe i forenzičare, povjesničare i lingviste. Tako velik interes izazvao je rad A. Wilson s Kalifornijskog sveučilišta. Godine 1987. objavio je rezultate usporedne analize DNK mitohondrija uzete od 147 predstavnika različitih etničkih skupina svih ljudskih rasa nastanjenih na pet kontinenata. Vrstom, mjestom i brojem pojedinačnih mutacija utvrđeno je da je sva mitohondrijska DNA nastala iz iste sekvence nukleotida predaka divergencijom. U pseudoznanstvenom tisku ovaj je zaključak tumačen na krajnje pojednostavljen način - cijelo je čovječanstvo potjecalo od jedne žene po imenu mitohondrijska Eva (a kćeri i sinovi mitohondrije primaju samo od svoje majke), koja je živjela u sjeveroistočnoj Africi prije oko 200 tisuća godina . Nakon još 10 godina bilo je moguće dešifrirati fragment mitohondrijske DNK izolirane iz ostataka neandertalca, te procijeniti životni vijek posljednjeg zajedničkog pretka čovjeka i neandertalca prije 500 tisuća godina.
Danas se ljudska mitohondrijska genetika intenzivno razvija kako u populaciji tako i u medicinskom aspektu. Uspostavljena je veza između niza teških nasljednih bolesti i defekata u mitohondrijskoj DNA. Genetske promjene povezane sa starenjem najizraženije su u mitohondrijima. Što je mitohondrijski genom, koji se po veličini, obliku i genetskom kapacitetu razlikuje kod ljudi i drugih životinja od biljaka, gljiva i protozoa? Kako funkcionira mitohondrijski genom i kako se pojavio u različitim svojtama? O tome će biti riječi u našem članku.
Mitohondrije se nazivaju elektranama stanice. Osim vanjske glatke membrane, imaju i unutarnju membranu koja tvori brojne nabore - kristale. Oni imaju ugrađene proteinske komponente respiratornog lanca - enzime koji sudjeluju u pretvaranju energije kemijskih veza oksidiranih hranjivih tvari u energiju molekula adenozin trifosforne kiseline (ATP). S ovom "konvertibilnom valutom" ćelija plaća sve svoje energetske potrebe. U stanicama zelenih biljaka osim mitohondrija postoje i druge energetske postaje - kloroplasti. Rade na "solarnim ćelijama", ali također stvaraju ATP iz ADP -a i fosfata. Poput mitohondrija, kloroplasti - autonomno reproducirajuće organele - također imaju dvije membrane i sadrže DNK.
Osim DNK, mitohondrijski matriks sadrži vlastite ribosome, koji se po mnogim karakteristikama razlikuju od eukariotskih ribosoma koji se nalaze na membranama endoplazmatskog retikuluma. Međutim, na ribosomima mitohondrija ne nastaje više od 5% svih proteina koji čine njihov sastav. Većina proteina koji čine strukturne i funkcionalne komponente mitohondrija kodirani su nuklearnim genomom, sintetizirani na ribosomima endoplazmatskog retikuluma i transportirani njegovim kanalima do mjesta okupljanja. Tako su mitohondriji rezultat zajedničkih napora dva genoma i dva stroja za transkripciju i translaciju. Neke podjedinice enzima mitohondrijskog respiratornog lanca sastavljene su od različitih polipeptida, od kojih su neki kodirani nuklearnim, a neki mitohondrijskim genomom. Na primjer, ključni enzim oksidativne fosforilacije, citokrom c oksidaza, u kvascu sastoji se od tri podjedinice kodirane i sintetizirane u mitohondrijima, te četiri podjedinice kodirane u staničnoj jezgri i sintetizirane u citoplazmi. Ekspresiju većine mitohondrijskih gena kontroliraju specifični geni u jezgri.
Veličine i oblici mitohondrijskih genoma
Do danas je pročitano više od 100 različitih mitohondrijskih genoma. Skup i broj njihovih gena u mitohondrijskoj DNA, za koje je nukleotidna sekvenca u potpunosti određena, uvelike se razlikuju kod različitih vrsta životinja, biljaka, gljiva i protozoa. Najveći broj gena pronađenih u mitohondrijskom genomu flagelatnih protozoa Rectinomonas americana- 97 gena, uključujući sve gene koji kodiraju proteine koji se nalaze u mtDNA drugih organizama. U većine viših životinja mitohondrijski genom sadrži 37 gena: 13 za proteine respiratornog lanca, 22 za tRNA i dva za rRNA (za veliku 16S rRNA podjedinicu ribosoma i za malu 12S rRNA). U biljkama i protozoama, za razliku od životinja i većine gljiva, neki proteini koji čine ribosome ovih organela kodirani su u mitohondrijskom genomu. Ključni enzimi sinteze predložaka polinukleotida, poput DNK polimeraze (replicirajuća mitohondrijska DNA) i RNA polimeraze (koja prepisuje mitohondrijski genom), kodirani su u jezgri i sintetizirani na citoplazmatskim ribosomima. Ova činjenica ukazuje na relativnost mitohondrijske autonomije u složenoj hijerarhiji eukariotske stanice.
Genomi mitohondrija različitih vrsta razlikuju se ne samo po skupu gena, redoslijedu njihovog smještaja i ekspresije, već i po veličini i obliku DNK. Velika većina danas opisanih mitohondrijskih genoma su kružne, super namotane dvolančane molekule DNA. U nekim biljkama, uz kružne oblike, postoje i linearni, a u nekih protozoa, na primjer, cilijata, u mitohondrijima se nalazi samo linearna DNA.
Obično svaki mitohondrij sadrži nekoliko kopija svog genoma. Dakle, u stanicama ljudske jetre postoji oko 2 tisuće mitohondrija, a u svakoj od njih ima 10 identičnih genoma. U mišjim fibroblastima postoji 500 mitohondrija koji sadrže dva genoma, te u stanicama kvasca S.cerevisiae- do 22 mitohondrija s četiri genoma.
Mitohondrijski genom biljaka u pravilu se sastoji od nekoliko molekula različitih veličina. Jedan od njih, "glavni kromosom", sadrži većinu gena, dok kraći kružni oblici, koji su u dinamičkoj ravnoteži međusobno i s glavnim kromosomom, nastaju kao posljedica unutar- i međumolekulske rekombinacije zbog prisutnost ponovljenih sekvenci (slika 1).
Sl. 1. Shema nastanka kružnih molekula DNK različitih veličina u biljnim mitohondrijima.
Rekombinacija se događa na ponovljenim područjima (označena plavom bojom).

Slika 2. Shema stvaranja linearnih (A), kružnih (B), lančanih (C) oligomera mtDNA.
ori - regija početka replikacije DNK.
Veličina genoma mitohondrija različitih organizama kreće se od manje od 6 tisuća parova baza u plazmodiju malarije (uz dva gena rRNA, sadrži samo tri gena koji kodiraju proteine) do stotina tisuća parova baza u kopnenim biljkama (na primjer, u Arabidopsis thaliana iz obitelji krstašica 366924 parova baza). Istodobno, 7-8 puta se razlikuju u veličini mtDNA viših biljaka čak i unutar iste obitelji. Duljina mtDNA kralježnjaka neznatno se razlikuje: kod ljudi - 16.569 parova baza, kod svinja - 16350, kod dupina - 16330, kod kandži žabe Xenopus laevis- 17533, u šarana- 16400. Ovi su genomi također slični po lokalizaciji gena, od kojih se većina nalazi s kraja na kraj; u nekim slučajevima čak se i preklapaju, obično za jedan nukleotid, tako da je posljednji nukleotid jednog gena prvi u sljedećem. Za razliku od kralježnjaka, u biljkama, gljivama i protozoama, mtDNA sadrže do 80% nekodirajućih sekvenci. Redoslijed gena u genomima mitohondrija razlikuje se od vrste do vrste.
Visoka koncentracija reaktivnih vrsta kisika u mitohondrijima i slab sustav popravka povećavaju učestalost mutacija mtDNA za red veličine u usporedbi s nuklearnom. Radikali kisika uzrokuju specifične supstitucije C® T (deaminacija citozina) i G® T (oksidativno oštećenje gvanina), uslijed čega je vjerojatno mtDNA bogata AT-parovima. Osim toga, sve mtDNA imaju zanimljivo svojstvo - nisu metilirane, za razliku od nuklearne i prokariotske DNA. Poznato je da je metilacija (privremena kemijska modifikacija nukleotidne sekvence bez narušavanja kodirajuće funkcije DNA) jedan od mehanizama programirane inaktivacije gena.
Replikacija i transkripcija mitohondrijske DNA sisavaca
U većine životinja komplementarne niti u mtDNA značajno se razlikuju po specifičnoj težini, budući da sadrže nejednaku količinu "teških" purinskih i "lakih" pirimidinskih nukleotida. Tako se zovu - H (teški - teški) i L (lagani - lagani) lanac. Na početku replikacije molekule mtDNA nastaje takozvana D-petlja (iz engleske petlje pomaka). Ova se struktura, vidljiva pod elektronskim mikroskopom, sastoji od dvolančanih i jednolančanih (uvučeni dio H-lanca) presjeka. Dvolančanu regiju tvori dio L-lanca i komplementarni novosintetizirani fragment DNA od 450-650 (ovisno o vrsti organizma) nukleotida u duljini, koji ima prajmer ribonukleotida na 5'-kraju, što odgovara početnoj točki sinteze H-lanca (ori H) .L-lanac počinje tek kada kćer H-lanac dosegne točku ori L. To je zbog činjenice da je područje početka replikacije L-lanca dostupan je enzimima sinteze DNA samo u jednolančanom stanju, pa stoga samo u ispletenoj dvostrukoj spirali tijekom sinteze H. Tako se kćeri lanci mtDNA sintetiziraju kontinuirano i asinkrono (slika 3 ).

Slika 3. Shema replikacije mtDNA sisavaca.
Prvo se formira D-petlja, zatim se sintetizira kćer H-lanac,
tada počinje sinteza kćeri L-lanca.
U mitohondrijima ukupni broj molekula D-petlje značajno premašuje broj molekula koje se potpuno repliciraju. To je zbog činjenice da D -petlja ima dodatne funkcije - vezanje mtDNA na unutarnju membranu i početak transkripcije, budući da su promotori transkripcije oba lanca DNA lokalizirani u ovoj regiji.
Za razliku od većine eukariotskih gena, koji se transkribiraju neovisno jedan o drugom, svaki od mtDNA lanaca sisavaca prepisuje se u jednu molekulu RNA koja počinje u regiji ori H. Uz ove dvije duge molekule RNA komplementarne H- i L-lancima , više kratkih dijelova H-lanca koji počinju na istoj točki i završavaju na 3 'kraju gena 16S rRNA (slika 4). Postoji 10 puta više takvih kratkih transkripata nego dugih. Kao rezultat sazrijevanja (obrada), od njih se formira 12S rRNA i 16S rRNA, koje sudjeluju u stvaranju mitohondrijskih ribosoma, kao i fenilalanin i valin tRNA Preostale tRNA se izrezuju iz dugih transkripata i stvaraju se prevedene mRNA, do 3 ' krajevi na koje su pričvršćene poliadenilne sekvence. 5 "krajevi ovih mRNA nisu prekriveni, što je neuobičajeno za eukariote. Ne dolazi do spajanja jer niti jedan od mitohondrijskih gena sisavaca ne sadrži introne.

Slika 4. Transkripcija humane mtDNA koja sadrži 37 gena. Svi se transkripti počinju sintetizirati u regiji ori H. Ribosomske RNA se izrezuju iz transkripata dugog i kratkog H lanca. tRNA i mRNA nastaju kao rezultat obrade iz transkripata oba lanca DNA. Geni tRNA prikazani su svijetlozeleno.Mitohondrijski genom iznenađuje
Unatoč činjenici da genomi mitohondrija sisavaca i kvasca sadrže približno isti broj gena, veličina genoma kvasca je 4-5 puta veća - oko 80 tisuća parova baza. Iako su mdDNA kodirajuće sekvence kvasca vrlo homologne onima u ljudi, mRNA kvasca dodatno imaju 5 "vodeću i 3" nekodirajuću regiju, poput većine nuklearnih mRNA. Brojni geni također sadrže introne. Tako okvirni gen koji kodira citokrom oksidazu b sadrži dva introna. Kopija većine prvog introna autokatalitički se izrezuje iz primarnog RNA transkripta (bez sudjelovanja bilo kakvih proteina). Preostala RNA služi kao predložak za stvaranje enzima maturaze za spajanje. Dio njegove aminokiselinske sekvence kodiran je u preostalim kopijama introna. Maturaza ih izrezuje, uništavajući vlastitu mRNA, kopije egzona se spajaju i tvori se mRNA za citokrom oksidazu b (slika 5). Otkriće takvog fenomena natjeralo nas je da preispitamo koncept introna kao "ništa što kodira sekvence". 
Slika 5. Obrada (sazrijevanje) mRNA citokrom oksidaze b u mitohondrijima kvasca.
U prvoj fazi spajanja nastaje mRNA, prema kojoj se sintetizira maturaza,
potrebno za drugu fazu spajanja.
Prilikom proučavanja ekspresije mitohondrijskih gena Trypanosoma brucei otkrio iznenađujuće odstupanje od jednog od osnovnih aksioma molekularne biologije koji kaže da se slijed nukleotida u mRNA točno podudara s onim u kodirajućim regijama DNA. Pokazalo se da se mRNA jedne od podjedinica citokrom c oksidaze uređuje, t.j. nakon transkripcije mijenja se njegova primarna struktura - umetnuta su četiri uracila. Kao rezultat, nastaje nova mRNA, koja služi kao predložak za sintezu dodatne podjedinice enzima, čija aminokiselinska sekvenca nema ništa zajedničko sa sekvencom kodiranom neuređenom mRNA (vidi tablicu).

Uređivanje RNA, prvi put otkriveno u mitohondrijima tripanosoma, rašireno je u kloroplastima i mitohondrijima viših biljaka. Nađen je i u somatskim stanicama sisavaca, na primjer, u ljudskom crijevnom epitelu, uređuje se mRNA gena apolipoproteina.
Najveće iznenađenje znanstvenicima predstavljale su mitohondrije 1979. Do tada se vjerovalo da je genetski kod univerzalan i da iste trojke kodiraju iste aminokiseline u bakterijama, virusima, gljivama, biljkama i životinjama. Engleski istraživač Burrell usporedio je strukturu jednog od mitohondrijskih gena teleta sa aminokiselinskom sekvencom u podjedinici citokrom oksidaze koju kodira ovaj gen. Pokazalo se da se genetski kod mitohondrija kod goveda (kao i kod ljudi) ne razlikuje samo od univerzalnog, već je “idealan”, tj. poštuje sljedeće pravilo: "ako dva kodona imaju dva identična nukleotida, a treći nukleotidi pripadaju istoj klasi (purin - A, G ili pirimidin - Y, C), tada kodiraju istu aminokiselinu". U univerzalnom kodu postoje dvije iznimke od ovog pravila: AUA triplet kodira izoleucin, a AUG kodon - metionin, dok u idealnom mitohondrijskom kodu oba ova tripleta kodiraju metionin; UGG triplet kodira samo triptofan, a UGA triplet kodira stop kodon. U univerzalnom kodu oba odstupanja odnose se na temeljne trenutke sinteze proteina: AUG kodon je inicijator, a UGA stop kodon zaustavlja sintezu polipeptida. Idealni kod nije svojstven svim opisanim mitohondrijima, ali niti jedan od njih nema univerzalni kod. Možemo reći da mitohondriji govore različitim jezicima, ali nikada ne govore jezikom jezgre.
Kao što je već spomenuto, u mitohondrijskom genomu kralježnjaka postoje 22 gena tRNA. Kako takav nepotpuni skup služi svih 60 kodona za aminokiseline (u idealnom kodu od 64 trojke postoje četiri stop kodona, u univerzalnom - tri)? Činjenica je da se tijekom sinteze proteina u mitohondrijima interakcije kodon -antikodon pojednostavljuju - dva od tri antikodon nukleotida koriste se za prepoznavanje. Dakle, jedna tRNA prepoznaje sva četiri člana obitelji kodona, koji se razlikuju samo po trećem nukleotidu. Na primjer, leucinska tRNA s GAU antikodonom stoji na ribosomu nasuprot kodona CUU, CUC, CUA i CUG, osiguravajući ugradnju leucina bez grešaka u polipeptidni lanac. Druga dva leucinska kodona, UUA i UUG, prepoznaju tRNA s antikodonskim AAU. Ukupno osam različitih molekula tRNA prepoznaje osam obitelji od po četiri kodona, a 14 tRNA prepoznaje različite parove kodona, od kojih svaka šifrira jednu aminokiselinu.
Važno je da su enzimi aminoacil tRNA sintetaze odgovorni za vezanje aminokiselina na odgovarajuću tRNA mitohondrija kodirani u jezgri stanice i sintetizirani na ribosomima endoplazmatskog retikuluma. Tako su u kralježnjaka sve proteinske komponente sinteze mitohondrijskog polipeptida kodirane u jezgri. Istodobno, sintezu proteina u mitohondrijima ne potiskuje cikloheksimid, koji blokira rad eukariotskih ribosoma, ali je osjetljiv na antibiotike eritromicin i kloramfenikol, koji inhibiraju sintezu proteina u bakterijama. Ova činjenica služi kao jedan od argumenata u prilog nastanku mitohondrija iz aerobnih bakterija tijekom simbiotskog stvaranja eukariotskih stanica.
Simbiotska teorija o postanku mitohondrija
Hipotezu o podrijetlu mitohondrija i biljnih plastida iz unutarstaničnih bakterija-endosimbiona izrazio je R. Altman još 1890. Tijekom stoljeća brzog razvoja biokemije, citologije, genetike i molekularne biologije koja se pojavila prije pola stoljeća, hipoteza je porasla u teoriju koja se temelji na velikoj količini činjeničnog materijala. Njegova je bit sljedeća: pojavom fotosintetskih bakterija kisik se nakupio u Zemljinoj atmosferi - nusprodukt njihova metabolizma. S povećanjem njegove koncentracije život anaerobnih heterotrofa postao je teži, a neki od njih su, radi dobivanja energije, prešli iz anoksične fermentacije u oksidativnu fosforilaciju. Takvi aerobni heterotrofi mogli bi, s većom učinkovitošću od anaerobnih bakterija, razgraditi organsku tvar nastalu fotosintezom. Neki od slobodnih aeroba uhvaćeni su anaerobima, ali nisu "probavljeni", već su sačuvani kao energetske stanice, mitohondriji. Mitohondrije ne treba promatrati kao robove koji su zarobljeni radi opskrbe molekula ATP stanicama koje ne mogu disati. Umjesto toga, oni su "stvorenja" koja su, još u proterozoiku, pronašla najbolja skloništa za sebe i svoje potomke, gdje možete uložiti najmanje truda, a da ne riskirate da vas pojedu.
Brojne činjenice govore u prilog simbiotičkoj teoriji:
- veličina i oblik mitohondrija i slobodnih aerobnih bakterija su isti; obje sadrže kružne molekule DNA koje nisu povezane s histonima (za razliku od linearne nuklearne DNA);Postoji ideja da su različita carstva eukariota imala različite pretke, a endosimbioza bakterija je nastala u različitim fazama evolucije živih organizama. O tome svjedoče i razlike u strukturi mitohondrijskih genoma protozoa, gljiva, biljaka i viših životinja. No u svim je slučajevima većina gena iz promitohondrija ušla u jezgru, vjerojatno uz pomoć pokretnih genetskih elemenata. Kada se dio genoma jednog od simbionta uključi u genom drugog, integracija simbionta postaje nepovratna.U smislu nukleotidnih sekvenci, ribosomske i transportne RNA mitohondrija razlikuju se od nuklearnih, dok pokazuju iznenađujuće sličnosti sa sličnim molekulama nekih aerobnih gram-negativnih eubakterija;
Mitohondrijske RNA polimeraze, iako kodirane u staničnoj jezgri, inhibirane su rifampicinom, poput bakterijskih, a eukariotske RNA polimeraze su neosjetljive na ovaj antibiotik;
Sintezu proteina u mitohondrijima i bakterijama potiskuju isti antibiotici koji ne utječu na ribosome eukariota;
Sastav lipida unutarnje membrane mitohondrija i bakterijske plazmaleme sličan je, ali se vrlo razlikuje od sastava vanjske membrane mitohondrija, koja je homologna drugim membranama eukariotskih stanica;
Cristae, nastale unutarnjom mitohondrijskom membranom, evolucijski su analozi mezosomalnih membrana mnogih prokariota;
Do sada su organizmi preživjeli koji oponašaju srednje oblike na putu stvaranja mitohondrija iz bakterija (primitivna ameba Pelomyxa nema mitohondrije, ali uvijek sadrži endosimbiotske bakterije).
Novi genom može stvoriti metaboličke putove koji vode do stvaranja korisnih proizvoda koje ne može sintetizirati niti jedan od partnera pojedinačno. Dakle, sinteza steroidnih hormona stanicama kore nadbubrežne žlijezde složen je lanac reakcija, od kojih se neke javljaju u mitohondrijima, a neke u endoplazmatskom retikulumu. Hvatanjem gena promitohondrija jezgra je mogla pouzdano kontrolirati funkcije simbionta. Jezgra kodira sve proteine i sintezu lipida vanjske membrane mitohondrija, većinu proteina matriksa i unutarnje membrane organela. Ono što je najvažnije, jezgra kodira enzime za replikaciju, transkripciju i translaciju mtDNA, kontrolirajući tako rast i reprodukciju mitohondrija. Stopa rasta simbiotskih partnera trebala bi biti približno ista. Ako domaćin raste brže, onda će se sa svakom generacijom broj simbionta po pojedincu smanjivati, a na kraju će se pojaviti potomci bez mitohondrija. Znamo da svaka stanica spolno reproducirajućeg organizma sadrži mnogo mitohondrija koji repliciraju njihovu DNK u intervalu između podjela domaćina. Time se osigurava da će svaka od stanica kćeri primiti barem jednu kopiju mitohondrijskog genoma.
Citoplazmatsko nasljeđivanje
Osim što kodira ključne komponente respiratornog lanca i vlastiti aparat za sintezu proteina, mitohondrijski genom u nekim slučajevima sudjeluje u stvaranju nekih morfoloških i fizioloških karakteristika. Ove značajke uključuju sindrom NCS-a (ne-kromosomske pruge, nekromosomski kodirane pjegavosti lista), karakterističan za brojne više biljne vrste, i citoplazmatsku mušku sterilnost (CMS), što dovodi do poremećaja normalnog razvoja peludi. Očitovanje obje značajke posljedica je promjena u strukturi mtDNA. U CMS -u se opaža preuređivanje mitohondrijskih genoma kao rezultat rekombinacijskih događaja koji dovode do delecija, duplikacija, inverzija ili umetanja određenih nukleotidnih sekvenci ili cijelih gena. Takve promjene mogu uzrokovati ne samo oštećenje postojećih gena, već i pojavu novih radnih gena.
Citoplazmatska nasljednost, za razliku od nuklearne, ne poštuje Mendelove zakone. To je zbog činjenice da u višim životinjama i biljkama gamete različitih spolova sadrže neusporedive količine mitohondrija. Dakle, u jajetu miša ima 90 tisuća mitohondrija, a u spermi - samo četiri. Očito, u oplođenom jajašcu mitohondriji su pretežno ili samo od ženke, t.j. nasljedstvo svih mitohondrijskih gena je majčino. Genetska analiza citoplazmatske nasljednosti je teška zbog nuklearno-citoplazmatskih interakcija. U slučaju citoplazmatske muške sterilnosti, mutirani mitohondrijski genom stupa u interakciju s određenim nuklearnim genima, čiji su recesivni aleli neophodni za razvoj svojstva. Dominantni aleli ovih gena, i u homo- i u heterozigotnim stanjima, vraćaju plodnost biljaka bez obzira na stanje mitohondrijskog genoma.
Proučavanje mitohondrijskih genoma, njihova evolucija prema posebnim zakonima populacijske genetike, odnos između nuklearnog i mitohondrijskog genetskog sustava, potrebno je za razumijevanje složene hijerarhijske organizacije eukariotske stanice i organizma u cjelini.
Određene nasljedne bolesti i starenje ljudi povezani su s određenim mutacijama u mitohondrijskoj DNA ili u nuklearnim genima koji kontroliraju mitohondrije. Akumuliraju se podaci o uključenosti defekata mtDNA u karcinogenezu. Stoga mitohondriji mogu biti meta kemoterapije raka. Postoje činjenice o bliskoj interakciji nuklearnog i mitohondrijskog genoma u razvoju niza ljudskih patologija. Više je delecija mtDNA pronađeno u pacijenata s teškom mišićnom slabošću, ataksijom, gluhoćom, mentalnom retardacijom, naslijeđenom na autosomno dominantan način. Utvrđen spolni dimorfizam u kliničkim manifestacijama koronarne bolesti srca, koji je najvjerojatnije posljedica majčinog učinka - citoplazmatske nasljednosti. Razvoj genske terapije nudi nadu za ispravljanje nedostataka u mitohondrijskim genomima u doglednoj budućnosti.
Ovaj rad je podržan od strane Ruske zaklade za temeljna istraživanja. Projekt 01-04-48971.
Autor je zahvalan diplomskom studentu M.K. Ivanovu, koji je stvorio brojke za članak.
Književnost
1. Yankovsky N.K., Borinskaya S.A. Naša povijest zabilježena u DNK // Priroda. 2001. broj 6. S.10-18. 2. Minchenko A.G., Dudareva N.A. Mitohondrijski genom. Novosibirsk, 1990. 3. Gvozdev V.A.// Soros. obrazovan. zhurn. 1999. broj 10. S.11-17. 4. Margelis L. Uloga simbioze u evoluciji stanica. M., 1983. godine. 5. Skulačev V.P.// Soros. obrazovan. zhurn. 1998. broj 8. S.2-7. 6. Igamberdiev A.U.// Soros. obrazovan. zhurn. 2000. Broj 1. S.32-36.
Mitohondrijska DNA smještena u matrici zatvorena je kružna dvolančana molekula u ljudskim stanicama veličine 16569 parova nukleotida, što je otprilike 10 5 puta manje od DNA lokalizirane u jezgri. Općenito, mitohondrijska DNA kodira 2 rRNA, 22 tRNA i 13 podjedinica enzima respiratornog lanca, što nije više od polovice proteina koji se nalaze u njoj. Konkretno, pod kontrolom mitohondralnog genoma kodirano je sedam podjedinica ATP sintetaze, tri podjedinice citokrom oksidaze i jedna podjedinica ubikinol-citokroma. s-reduktaze. U ovom slučaju, svi proteini, osim jedne, dvije ribosomske i šest tRNA, transkribirani su iz težeg (vanjskog) lanca DNA, a 14 drugih tRNA i jedan protein prepisani su iz lakšeg (unutarnjeg) lanca.
U tom kontekstu, genom biljnih mitohondrija mnogo je veći i može doseći 370 000 parova nukleotida, što je oko 20 puta veće od gore opisanog genoma mitohondrija čovjeka. Ovdje je broj gena također oko 7 puta veći, što je popraćeno pojavom u biljnim mitohondrijima dodatnih transportnih putova elektrona koji nisu povezani sa sintezom ATP -a.
Mitohondrijska DNA replicira se u interfazi, što je djelomično sinkronizirano s replikacijom DNA u jezgri. Tijekom staničnog ciklusa mitohondriji se sužavanjem dijele na dva dijela, čije formiranje počinje prstenastim žlijebom na unutarnjoj membrani mitohondrija. Detaljno proučavanje nukleotidne sekvence mitohondrijskog genoma omogućilo je ustanoviti da su u mitohondrijima životinja i gljiva česta odstupanja od univerzalnog genetskog koda. Tako u ljudskim mitohondrijima kodon TAT umjesto izoleucina u standardnom kodu kodira aminokiselinu metionin, kodoni TCT i TCC, koji obično kodiraju arginin, su stop kodoni, a AST kodon koji je stop kodon u standardnom kodu , kodira aminokiselinu metionin. Što se tiče biljnih mitohondrija, čini se da koriste univerzalni genetski kod. Druga značajka mitohondrija je prepoznavanje kodona tRNA, što se sastoji u činjenici da jedna takva molekula može prepoznati ne jedan, već tri ili četiri kodona odjednom. Ova značajka smanjuje važnost trećeg nukleotida u kodonu i dovodi do činjenice da mitohondrije zahtijevaju manji broj tipova tRNA. U ovom slučaju dovoljne su samo 22 različite tRNA.
Budući da ima svoj genetski aparat, mitohondrije imaju i vlastiti sustav za sintezu proteina, čija je značajka u stanicama životinja i gljiva vrlo mali ribosomi koje karakterizira koeficijent sedimentacije 55S, koji je čak niži od onog iz 70-ih ribosoma prokariotski tip. U ovom slučaju, dvije velike ribosomske RNK su također manje nego kod prokariota, a mala rRNA potpuno nedostaje. U biljnim mitohondrijima, naprotiv, ribosomi su po veličini i građi sličniji prokariotskim.
Svojstva i funkcije DNK.
DNK ili deoksiribonukleinska kiselina glavni je nasljedni materijal prisutan u svim stanicama tijela i uglavnom uključuje plavi pečat staničnih funkcija, rasta, reprodukcije i smrti. Strukturu DNA koja se naziva dvolančana spiralna struktura prvi su opisali Watson i Crick 1953. godine.
Od tada je postignut ogroman napredak u sintezi, sekvenciranju i manipulaciji DNK. DNK se ovih dana može virtualizirati ili analizirati na sitnice, pa čak i ubaciti gene da izazovu promjene u funkciji i strukturi DNA.
Glavna svrha nasljednog materijala je pohrana nasljednih informacija na temelju kojih se formira fenotip. Većina karakteristika i svojstava organizma posljedica je sinteze bjelančevina koje obavljaju različite funkcije. Dakle, podaci o građi izrazito različitih molekula bjelančevina čija specifičnost ovisi i o kvalitativnom i kvantitativnom sastavu aminokiselina kao i po redoslijedu njihovog rasporeda u peptidnom lancu, moraju se zabilježiti u nasljednom materijalu. Posljedično, aminokiselinski sastav proteina mora biti kodiran u molekulama nukleinske kiseline.
Još u ranim 50 -im godinama predložena je metoda za bilježenje genetskih informacija, u kojoj bi se kodiranje pojedinih aminokiselina u molekuli proteina trebalo provesti korištenjem određenih kombinacija četiri različita nukleotida u molekuli DNA. Za šifriranje više od 20 aminokiselina potreban broj kombinacija daje samo tripletni kôd, odnosno kôd koji uključuje tri susjedna nukleotida. U ovom slučaju, broj kombinacija četiri dušične baze, svaka po tri je 41 = 64. Pretpostavka o trojstvu genetskog koda kasnije je eksperimentalno potvrđena, a za razdoblje od 1961. do 1964. pronađena je šifra, uz pomoć od kojih je redoslijed aminokiselina zapisan u molekulama nukleinske kiseline u peptidu.
Sa stola. 6 da od 64 tripleta, 61 triplet kodira jednu ili drugu aminokiselinu, a pojedinačne aminokiseline su šifrirane s više od jednog tripleta ili kodona (fenilalanin, leucin, valin, serin itd.). Nekoliko trojki ne kodira aminokiseline, a njihove su funkcije povezane s označavanjem krajnjeg područja proteinske molekule.
Čitanje informacija zabilježenih u molekuli nukleinske kiseline provodi se uzastopno, zajedno s kodonom, tako da je svaki nukleotid dio samo jednog tripleta.
Proučavanje genetskog koda u živim organizmima s različitim razinama organizacije pokazalo je svestranost ovog mehanizma za bilježenje informacija u živoj prirodi.
Tako su istraživanja sredinom 20. stoljeća otkrila mehanizam bilježenja nasljednih informacija u molekulama nukleinske kiseline pomoću biološkog koda, koji se odlikuje sljedećim svojstvima: a) triplet - aminokiseline su šifrirane trojkama nukleotida - kodona; b) specifičnost - svaki triplet kodira samo određenu aminokiselinu; c) univerzalnost - u svim živim organizmima kodiranje istih aminokiselina provode isti kodoni; d) degeneracija - mnoge aminokiseline su šifrirane s više od jednog tripleta; e) ne preklapaju se - čitanje informacija vrši se uzastopno trostruko po tripletno: AAGCTCagCCCTT.
Osim bilježenja i pohranjivanja bioloških informacija, funkcija nasljednog materijala je i njegova reprodukcija i prijenos na novu generaciju u procesu razmnožavanja stanica i organizama. Ovu funkciju nasljednog materijala molekule DNA provode u procesu njezine reduplikacije, tj. Apsolutno točne reprodukcije strukture, zahvaljujući provedbi načela komplementarnosti (vidi 2.1).
Konačno, treća funkcija nasljednog materijala, predstavljenog molekulama DNA, jest pružanje specifičnih procesa tijekom implementacije informacija sadržanih u njemu. Ova se funkcija provodi uz sudjelovanje različitih vrsta RNA, koje osiguravaju proces translacije, odnosno sastavljanje molekule proteina koja se javlja u citoplazmi na temelju informacija dobivenih iz jezgre (vidi 2.4). Tijekom ostvarivanja nasljednih informacija pohranjenih u obliku molekula DNA u kromosomima jezgre razlikuje se nekoliko faza.
1. Čitanje informacija iz molekule DNA u procesu sinteze mRNA - transkripcija, koja se provodi na jednom od lanaca dvostruke spirale DNA -kodogenog lanca prema načelu komplementarnosti (vidi 2.4).
2. Priprema produkta transkripcije za otpuštanje u citoplazmu - sazrijevanje mRNA.
3. Sastavljanje na ribosomima peptidnog lanca aminokiselina na temelju informacija zabilježenih u molekuli mRNA, uz sudjelovanje transportne tRNA - translacija (vidi 2.4).
4. Formiranje sekundarnih, tercijarnih i kvartarnih proteinskih struktura, što odgovara stvaranju funkcionalnog proteina (jednostavna značajka).
5. Formiranje složene osobine kao rezultat sudjelovanja produkata nekoliko gena (proteinski enzimi ili drugi proteini) u biokemijskim procesima.
Struktura dvostruke spirale DNA, koju zajedno drže samo vodikove veze, može se lako uništiti. Razbijanje vodikovih veza između polinukleotidnih DNA lanaca može se provesti u jako alkalnim otopinama (pri pH> 12,5) ili zagrijavanjem. Nakon toga se lanci DNA potpuno odvajaju. Taj se proces naziva DNK denaturacija ili taljenje.
Denaturacija mijenja neka fizička svojstva DNK, poput optičke gustoće. Dušične baze apsorbiraju svjetlost u ultraljubičastom području (s maksimalno blizu 260 nm). DNK apsorbira svjetlost gotovo 40% manje od mješavine slobodnih nukleotida istog sastava. Ta se pojava naziva hipokromni učinak, a nastaje zbog interakcije baza kada se nalaze u dvostrukoj spirali.
Svako odstupanje od dvolančanog stanja utječe na promjenu veličine ovog učinka, tj. dolazi do pomaka optičke gustoće prema vrijednosti karakterističnoj za slobodne baze. Tako se denaturacija DNA može promatrati promjenom njezine optičke gustoće.
Kada se DNK zagrije, prosječna temperatura raspona u kojem se odvajaju lanci DNK naziva se talište i označava kao T pl... U otopini T pl obično se nalazi u rasponu 85-95 ° C. Krivulja taljenja DNA uvijek ima isti oblik, ali njezin položaj na temperaturnoj ljestvici ovisi o sastavu baze i uvjetima denaturacije (slika 1). G-C parovi povezani s tri vodikove veze su vatrostalniji od AT parova koji imaju dvije vodikove veze, pa s povećanjem sadržaja G-C-nap vrijednost T pl povećava. DNK, 40% sastavljena od G-C (karakteristična za genom sisavaca), denaturira se pri T pl oko 87 ° C, dok DNA koja sadrži 60% G-C ima T pl
oko 95 ° C.
Na temperaturu denaturacije DNA (osim sastava baza) utječe ionska snaga otopine. Štoviše, što je veća koncentracija monovalentnih kationa, veći je T pl... T vrijednost pl također se uvelike mijenja kada se u otopinu DNA dodaju tvari poput formamida (amid mravlje kiseline HCONH2), koji
destabilizira vodikove veze. Njegova prisutnost omogućuje vam smanjenje T pl, do 40 ° C.
Proces denaturacije je reverzibilan. Pojava obnove strukture dvostruke spirale, temeljena na dva odvajanja komplementarnih niti, naziva se renaturacija DNA. Za renaturaciju je u pravilu dovoljno pritisnuti otopinu denaturirane DNA.
Renaturacija uključuje dva komplementarna niza koja su odvojena tijekom denaturacije. Međutim, sve komplementarne sekvence koje mogu oblikovati dvolančanu strukturu mogu se ponovno naseliti. Ako zajedno. jednolančana DNA koja potječe s različitih točaka se žari, tada se stvaranje dvolančane DNK strukture naziva hibridizacija.
Slične informacije.