Taimede peamiste organite funktsioonid. Kõrgemate taimede üldised omadused. Kõrgemate taimede erinevused madalamast. Taimede klassifikatsioon nende eluvormide jaoks, taimede kirjeldus
Taimede Kuningriik on selle ülevuse tõttu silmatorkav. Kui me läheme, mis tahes planeedi nurgas, saame kohtuda kõikjal taimemaailma esindajad. Isegi Arktika jää ei ole nende elupaiga erand. Mis see on taimi kuningriik? Selle esindajate tüübid on erinevad ja arvukad. Mis on taimi kuningriigi üldine omadus? Kuidas nad neid klassifitseerivad? Proovime välja selgitada.
Taimede Kuningriigi üldised omadused
Kõik elusorganismid võib jagada neljaks kuninglikuks: taimed, loomad, seente ja bakterite.
Taimete Kuningriigi tunnused on järgmised:
- on eukarüootid, st taimerakud sisaldavad tuumad;
- on autotroofid, mis moodustavad anorgaaniliste orgaaniliste ainete kujul fotosünteesi protsessis päikesevalguse energia tõttu;
- juhtida suhteliselt fikseeritud elustiili;
- piiramatu kasvu kogu elu jooksul;
- sisaldama plastiidide ja tselluloosi rakulisi kestad;
- vaba toitainetena kasutatakse tärklist;
- klorofülli olemasolu.
Taimede botaaniline klassifikatsioon
Taimede Kuningriik on jagatud kaheks rajatiseks:
- madalamad taimed;
- kõrgemad taimed.
Fitness "madalamad taimed"
Need rajatised hõlmavad vetikaid - kõige lihtsam struktuuri ja kõige vanemate taimede kohta. Veesoolade maailm on siiski väga mitmekesine ja arvukad.
Enamik neist elavad vees või vees. Aga seal on vetikad, mis kasvab mullas, puudel, kividel ja isegi jääl.
Vetikate keha on kihiline või pikk, millel pole juur ega võrseid. Vetikate puhul ei ole elundeid ega erinevaid kudesid, ainete (vee ja mineraalsoolade) imendumine kogu keha pinnaga.
"Madalamad taimed" koosneb üheteistkümnest vetikast osakonnast.
Isiku väärtus: hapnik; süüakse; agar-agari saamiseks; Kasutatakse väetistena.
Fitness "Kõrgemad taimed"
Kõrgeimad taimed hõlmavad organisme, millel on hästi väljendunud kangad, elundid (vegetatiivne: juur ja põgenemine, generatiivne) ja individuaalne areng (ontogenees), millest on jagatud embrüonaalseteks (mikroobeks) ja postitamisjärgse (postikoha) perioodideks.
Kõrgemad taimed on jagatud kaheks rühmaks: vaidlus ja seemned.
Spore taimed kehtivad vaidlusega. Vajaliku aretamiseks vee. Seemnete taimed kehtivad seemnetega. Aretusvette puhul ei ole vaja.
Spore taimed on jagatud järgmistesse osakondadesse:
- mossoid;
- playvoid;
- hang-kujuline;
- fern.
Seemned on jagatud järgmistesse osakondadesse:
- kaetud soolveega;
- lisatud.
Mõtle neid rohkem üksikasjalikumalt.
Osakond "Mohovoid"
Mossoid - need on madala rohttaimed, mille keha on jagatud varreteks ja lehtedeks, neil on mingi juured - rhizoid, mille funktsioon on absorbeerida vett ja määrata taim pinnasesse. Lisaks fotosünteetilisele ja peamisele kangale ei ole Mas'il teisi kudesid. Enamik MMS-i on mitmeaastased taimed ja kasvavad ainult niisketes kohtades. Mossoid - kõige iidsem ja kõige lihtsam rühm. Samal ajal on nad üsna mitmekesised ja paljud ja halvemad ainult kaetud taimeliikide koguses. Seal on umbes 25 tuhat liigi.
Mosoidid jagunevad kaheks klassiks - maksa- ja lehtköögiks.
Maksa - need on kõige iidsemad samblad. Nende keha on hargnenud lameda kiht. Enamasti troopikas. Maksa esindajad: MA Liikmednia ja Riccia.
Lubered Mossial on võrsed, mis koosnevad varredest ja lehtedest. Tüüpiline esindaja on Moss Kukushkini lina.
Makhovil võib olla seksuaalne ja võimsam reprodutseerimine. Casual võib olla nii vegetatiivne kui taimede tõugude osad varred, kihid või lehed ja vaidlus. Seksuaalse reproduktsiooniga moodustasid mossoidid spetsiaalseid organite, kus fikseeritud munad ja liikuvad spermatoosisa valmivad. Sperma vees liigub munadesse ja väetada neid. Siis taim kasvab kasti vaidlustega, mis on hajutatud pärast küpsemist ja levinud pikkade vahemaade.

Mossi eelistavad märgikohti, kuid nad kasvavad kõrbetes ja kivimites ning tundras, kuid neid ei leitud merede ja kõrge soolalahuse muldades lahtiste liivade ja liustikke.
Isiku väärtus: turba kasutatakse laialdaselt kütusena ja väetisena, samuti vaha, parafiini, värvide, paberi tootmiseks kasutatakse soojusisolatsioonimaterjalina konstruktsioonis.
Osakonnad "Playvoid", "Helds" ja "Fern"
Nendel kolme vaidluste taimede osakonnal on sarnane struktuur ja paljunemine, enamik neist kasvab varjulistes ja märgruumides. Nende taimede puitvormid on väga haruldased.
Ferns, lennukid ja hobused on iidsed taimed. 350 miljonit aastat tagasi nad esindasid suuri puid, see oli nad, kes moodustasid metsa planeedil, lisaks nad on praegu söehoiuste allikad.
Vähesed taimi liikide osakondade osakonnad Fern, need ja lennukikujulised, mis jõudis tänapäeva päeva, võib nimetada elus fossiilideks.
Väliselt erinevad plaatide, hobuste ja sõnajalate tüübid üksteisest erinevad. Kuid need on sisemise struktuuri ja paljunemise sarnased. Nad on raskem kui mossoidid (omama oma struktuuris rohkem kangaid), kuid see on lihtsam kui seemnetaimed. Viide vaidluste taimedele, nagu nad kõik moodustavad vaidlusi. Ka nende jaoks on võimalik nii seksuaalne kui ka mitte-reprodutseerimine.
Kõige iidsemad esindajad need eemaldamised on taimed. Tänapäeval okassaare metsades saate kohtuda Balavovoidiga.
Horsetails leidub põhjapoolkeral, nüüd neid esindavad ainult maitsetaimed. Horsetails võib leida metsade, soode ja niidud. Mängijate esindaja - valdkond, mis kasvab tavaliselt happelistes pinnas.
Kulunud - piisavalt suur rühm (umbes 12 tuhat liiki). Nende hulgas on rohi ja puud. See kasvab peaaegu kõikjal. Ferni esindajad - Ortrichnik ja Orlyaki esindajad tavalised.

Isiku tähendus: iidsed Fern-sarnased andsid meile kivisöe hoiused, mida kasutatakse kütusena ja väärtuslikke keemilise toorainena; Toidus kasutatakse mõningaid liike, kasutatakse väetistena kasutatavate ravimitena.
Osakond "Kogutud" (või "lill")
Lilletaimed on kõige paljude ja kõrge organiseeritud taimerühma. Seal on rohkem kui 300 tuhat liiki. See rühm on planeedi taimekaaruse peamine osa. Peaaegu kõik meid ümbritseva taimemaailma esindajad tavalises elus nii looduslikud kui aiataimed on kaetud sildade esindajad. Nende hulgas leiad kõik eluvormid: puud, põõsad ja maitsetaimed.
Kaetud taimede peamine erinevus on see, et nende seemned on kaetud Pestu käivitamisest moodustatud puuviljaga. Puuviljad on seemnete kaitse ja aitab kaasa nende levitamisele. Kaetud sillad moodustavad lilled - seksuaalse paljunemise organ. Kahekordne viljastamine on nende jaoks iseloomulik.

Lilletaimed valitsevad köögivilja-kaanega, mis on kõige kohandatud meie planeedi kaasaegsetele elutingimustele.
Isiku tähendus: kasutatakse toidus; eraldatud hapnikke keskkonda; kasutatakse ehitusmaterjalidena, kütusena; Kasutatakse meditsiinitööstuses, toiduainetes, parfümeeriatööstuses.
Osakonna "Hääletus"
Gamotical Taimed esindavad puud ja põõsad. Nende hulgas ei ole maitsetaimi. Enamikul gameerritel on lehed nõelate kujul (närimine). Hääletamisel on esile tõstetud suur okaspuude rühm.
Umbes 150 miljonit aastat tagasi valitsesid planeedi köögiviljakasvatusseadmed.

Tähendus isikule: moodustavad okasmetsad; eraldada suur hulk hapnikku; kasutatakse kütusena, ehitusmaterjalide, laevaehituse, mööbli tootmise; Kasutatakse meditsiinis toiduainetööstuses.
Erinevate taimede maailma, taimede nimed
Ülaltoodud klassifikatsioonil on jätkumised, osakonnad jagunevad klassideks, klassideks - tellimusteks, seejärel järgige perekonda, seejärel pereliike.
Taimede Kuningriik on tohutu ja mitmekesine, seega on tavaline kasutada kahe nimega taimede botaanilisi nimesid. Esimene sõna pealkiri tähendab taimede perekonda ja teine \u200b\u200bvaade. See on see, kuidas süstemaatika näeb meile kõigile kuulus Daisy apteek:
Kuningriik: taimed.
Osakond: lill.
Klass: Dichomotic.
Tellimus: Astrocolore.
Perekond: Astrovaya.
Varras: kummel.
Vaade: Daisy apteek.

Taimede klassifikatsioon nende eluvormide jaoks, taimede kirjeldus
Taimede Kuningriik klassifitseeritakse ka eluvormide kohaselt, st taimse organismi välimuse kohaselt.
- Puud - mitmeaastased taimed, millel on otsustavad õhuliinid ja üks pagasiruumi.
- Põõsad on ka mitmeaastased taimed, kellel on otsustavad õhuliinid, kuid erinevalt puudest ei ole neil selgelt väljendatud üks tünn ja hargnemine algab maa peal ja moodustub mitmed samaväärsed varred.
- Põõsad on sarnased põõsastega, kuid madalad - mitte kõrgemad kui 50 cm.
- Harticies on siiski sarnane põõsastega, aga need eristatakse asjaolu, et ainult pildiste alumised osad on kaunistatud ja ülemine surm.
- Liana - klammerdumine, ronimine ja lokkis varred.
- Sukulendid on mitmeaastased taimed lehed või varred, mis sisaldavad veevarustuse.
- Maitsetaimed - taimed rohelise, mahlakad ja soovimatud võrsed.
Looduslikud ja kultuurilised taimed
Käsi ja mees pani mitmesuguseid taimseid rahu ja tänapäeval võivad taimed jagada ka looduslikeks ja kultuurilisteks.

Garantii - taimed looduses, mis kasvavad, arendada ja rakendada ilma inimese abi.
Kultuuriettevõtted tekivad metsikult, kuid mis saadakse valiku, hübridiseerimise või geenitehnoloogia abil. Need on kõik aed taimed.
Teste
660-01. Spetsiaalsed õhuvarustuse asutuse taimed on
A) roheline leht
B) rootpode
C) lill
D) puuviljad
Vastus
660-02. Milline roll on juured mängima taime elus?
A) moodustavad orgaanilised ained anorgaanilisest
B) jahutatud taimed
C) vaba orgaanilised ained
D) Absorbeerib süsinikdioksiidi ja eriaine hapnikku
Vastus
660-03. Juure peamine funktsioon on
A) toitainete hüppeliselt
B) Mulla elektrijaamad
C) orgaaniliste ainete imendumine pinnast
D) orgaaniliste ainete oksüdeerimine
Vastus
660-04. Milline on lehte kõige olulisem roll taime elus?
A) tagab vee aurustumise
B) täidab võrdlusfunktsiooni
C) kasutatakse kaitseorganina
D) neelab vett ja mineraalsoolasid
Vastus
660-05. Mis seisundi vett võib taimse tõusta?
A) Vee aurustumise puudumisel
B) vee konstantse aurustamisega
C) ainult päevasel ajal
D) ainult suletud tolmuga
Vastus
660-06. Lehtede peamine roll taimede elus -
A) hingamine
B) Võimalik
C) fotosünteetiline
D) vegetatiivne reprodutseerimine
Vastus
660-07. Vee lehtede aurustamine aitab kaasa
A) mineraalsoolade liikumine taimes
B) orgaaniliste ainete lehtede pakkumine
C) süsinikdioksiidi imendumine kloroplastidega
D) orgaaniliste ainete moodustumise kiiruse suurenemine
Vastus
660-08. Peamine funktsioon varre on
A) õhuvõimsusega taimed
B) Veevarustus ja toitained
C) vesi ja toitained
D) vee aurustamine
Vastus
660-09. Mis järgmistest allpool on seade kuivadele elutingimustele?
A) Laged lehed
B) STOMPide komplekt
C) lihavad varred
D) pigistavad varred
Vastus
660-10. Mikuriza moodustavad seened saadakse taimede juurtest
A) vesi
B) antibiootikumid
C) mineraalsoolad
D) Orgaanilised ained
Vastus
660-11. Tremi roll taime elus koosneb
A) Taime tugevdamine pinnasesse
B) orgaaniliste ainete moodustumine
C) ainete liikumine taimele
D) vee imendumine ja mineraalsoolad
Maa biosfääris on vähemalt 5 (pigem 20 ja tõenäoliselt isegi 50) miljonit erinevat tüüpi elusolenditest. Taimestiku maailm on rikas ja mitmekesine (üle 500 tuhande liiki). Sageli jagab taime Kuningriik (Vegetabilia, Plantae või Phytobiota) kolm restrit: punased vetikad või vead (Rhodobionta), tõelised vetikad (Phycobionta) ja kõrgemate taimede või embrüobionta organismid (embrüobionta). Kaks esimest võimalust viitavad madalamatele taimedele (kihilised või tallomaatilised taimed - thallophyta). See rühm ühendab kõige lihtsalt organiseeritud taimed (üherakulise, koloonia ja multikullite), veekeskkonna elanike, eriti merede ja ookeanide elanike. Orgaanilise maailma mõnes kaasaegses süsteemis on sellel ajalooline tähtsus, kuna varem on eukarüootsed üherakulised, liikuvad vormid ja nüüd kõik madalamad taimed kuuluvad sageli Rubbs'i kuningriigis (Protista).
Kõrgemad taimed viitavad ka pildistamisele või lehtede sisaldusele (Cormobionta, Cormofyta); Telobionta telomofyta); maataimed.
Kõige kõrgemate taimede ees iseloomustavad järgmised põhifunktsioonid.
Enamasti maapinna elustiili. Kõrgemad taimed - õhukeskkonna elanikud. Pika evolutsioonilise protsessi ajal tekkisid uued liigid, mis on kohandatud elu tingimustes elu tingimustes. Vesiivormide olemasolu kõrgemate taimede seas (rdest, Elodea, vokandid ja teised) - nähtus on sekundaarne.
Väljapääs maapealse elupaikade tingimustele andis võimas tõuke kogu taime korraldamise ümberkorraldamisele. Maapealsetes tingimustes paranes taimevalgustus, mis aktiveeris fotosünteesiprotsessi, tõi kaasa assimiteatide suurenemise, taimede maht üldiselt põhjustas nende edasise morfoloogilise kaotamise vajaduse. Taimede vegetatiivne keha on jagatud kaheks osaks (jahvatatud ja maa-aluseks), mis täidab erinevaid funktsioone. Seega on kõrgemate taimede jaoks morfoloogiline lammutamist iseloomustavad kaks peamist vegetatiivset organitet: juur ja põgenemine. Varre ja lehed on põgenemise komponendid (nagu ühe tellimuse korpused). Kõik lehtede kõrgemad taimed kuuluvad morfoloogilise rühma - võrsed. Selle rühma kaasaegsete taimede seas domineerivad kõrge organiseeritud vormid, mis on kohandatud mitmesuguste sushi elutingimustega. Neid iseloomustab paljude peamiste organite ja nende komponentide (mugulad, puudusid, selg, vuntsid, kaalud, pirnid jt). Lihtsama organisatsiooniga on mitmeid primitiivseid esindajaid, kellel pole juured. Luminous sammal (Cukushkini lina jt) - esmane mittesiduvad taimed. Vett absorbeerimiseks moodustavad nad risooid. Bubble - teine \u200b\u200bkahetsusväärne taim (elab ülemise veekihid).
Mitte kõikidel kõrgeimal taimedel on ka vegetatiivsete organite üldosa. Maksa mays esitatakse sageli rohelise dikotoomiliselt hargneva plaadiga rohelise dikotoomia-hargneva plaadiga. Need moodustavad morfoloogilise rühma talamboomne või kihi taimed.
Kõrgemate taimede jaoks on iseloomulik keerulisem anatoomilise koe diferentseerumine. Õhutranspordi eristatakse paljude keskkonnategurite suurt immatkeerimist, nende suurema kontrastsuse. Maa tingimustes töötas taimed välja keerulise kate kangaste süsteemi. Kõige olulisem spetsialiseerunud kate kangast, ilma milleta sushi on võimatu, on epidermma. Evolutsioonikavas on see väga iidne päritolu. Esimese kuulsate maataimede (rhinofiit) keha oli kaetud epidermamiga, millel on tolmu gaasivahetuse ja transpiratsiooni reguleerimiseks. Tulevikus moodustati teisejärguline (periderma) ja tertsiaarne katmine (RITID) kangas.
Põhikeskkonnas sai enamik kõrgemaid taimi ka juhtivate, mehaaniliste, eri-, sisepiiride ja muude kudede kõige keerulisemate arengukompleksi.
Maa-aluste ja õhuliiride tavapärase toimimise jaoks on vaja vee, mineraalsete ja orgaaniliste ainete kiiret liikumist. See saavutatakse spetsiaalsete juhtivate kudede väljatöötamisega kõrgemate taimede kehas: ksümemid või puit, flora või loba. Upstream suunas, vastavalt ksüleen, vesi liigub mineraalsete ainetega, mis on selles lahustunud, imenduvad mulla juurtest ja orgaanilisest, toodetud juured ise. Kahanevas suunas, floem, assimilatsiooni tooted liigutatakse peamiselt süsivesikuid. Spetsiaalsed vee-juhtivad elemendid ksümemide on moodustatud - trahatide ja seejärel hingetoru või anumad. Ring ja spiraal tracheids ja laevad moodustavad protoksüüle, pooride, eri tüüpi külgmised poorid (trepikoda, vastupidine, regulaarne, mõnikord juhuslik) - metaktilise. Kõige iina tüüpi pooride tracheid on trepi. Need on iseloomulikud enamiku kõrgeimate spooride metateemile.
Vaidluste taimede seas on trepi laevad kättesaadavad mõnede sõnajalate metamiga (näiteks Orlyak) metamisomis. Laevad leidub ka kõrgelt organiseeritud hääletatud sekundaarse ksüleeni (Efedra, vastupidine ja Velvichia) sekundaarse ksüleen. Siiski on märgitud märgid ja gumpied laevad vähe ja peamine funktsionaalne koormus veetakse tracheids. Nad avastasid juba rinofiit.
Eeldatakse, et assimilatsiooni toodete transportimise elemendid taimemaailma arengu protsessis ilmus varem kui kastmine. Uriin tuvastas pikliku õhukese seinaga rakud plasmodaatide torude ja pooridega, mis võivad osaleda orgaaniliste lahenduste läbiviimisel. Lehekülaliste sammal, pikad õhukesed rakud teostatakse selle funktsiooni, mis on veidi pikemad otstes. Mitmed sellised rakud moodustavad pikisuunalise ühe rea. Ülejäänud kõrgeima vaidluse ja valija on iseloomulik süntoidrakkude moodustumisele. Ainult kaetud soolalahuses on moodustatud sõela torud moodustuvad segmendid, mis tekkisid sünovoidrakkudest. Nende konversioonide protsess on sarnane trahatide ümberkujundamise protsessiga laeva segmendis. Sünotoidtorud on tõhusamad kui sünovoid rakud teostavad fotosünteesi tooteid. Lisaks sellele on iga segmendis kaasas üks või mitu satelliitrakku.
Juhtivad elemendid, mehaanilised kangad ja parenhüümid on rühmitatud looduslikele kombinatsioonidesse - veresoonte kiud talad (radiaalne, kontsentriline, tagatis, Bicocollate). Kõige tavalisemad tagatised, avatud (dicotüatilised) ja suletud tüübid (ühe magamistoaga) taimed on kõige levinumad. Kesk-silinder - Stele, kõigepealt kõige lihtsamate algtoonide kujul. Tulevikus moodustuvad varre struktuuri suurendamise ja tüsistamise tõttu tekkinud SympoSose, näitleja, EUTELA ja ATACONTASK.
Kõrgematel taimedel on ka võimeline mehaaniliste kangate arendamine. Ajal elus veekeskkonnas, vajadus nende kudede oli väike, kuna vesi hästi toetas nende keha. Õhus, mille tihedus on mitu korda väiksem kui vesilahus, mehaanilised kuded pakuvad taimse staatilise (gravitatsiooni) ja dünaamilise (st kiiresti muutuva) koormuse vastupanuvõime (tuuleenergia, dušš, loomade kokkupuude jne). Protsessis areng, spetsialiseerunud mehaanilised kangad (Sclerchim ja kolledžum) tekkinud ja arendatud seoses progresseeruva lagemise ja suurenenud mass taimede.
Nagu elundid, ilmusid kangad ja ei arenenud kohe. Talombali kõrgemad taimed on välja töötatud juhtivast süsteemist. Mehaanilised kangad on ka halvasti arenenud. Väikesed suurused, polsterdatud märgades tingimustes, tagatakse nende taimede tugevus suures osas kestade elastsusest, mis moodustavad nende veega küllastunud rakud. Väljasurnud fossiilide uurimine avab kogu vahepealsete vormide read, mis näitavad teatud elundite ja kudede arendamise tekkimist ja etappe.
Kõrgemate taimede vaidlustel on keerulisem struktuur kui madalamad taimed (vetikad). Nad on statsionaarsed, ilma maitseta. Kõrgemate vaidluste lahendamine (mugh-kujuline, tavaline, külvamine, ferment ja psühho-kujuline) on kaetud mitmekihilise rakulise kestaga (Spidermiga). See koosneb kahest peamisest kihist: tahke väljas (exverse) ja trahvi sisemine (endositsioon). Tselluloosi sisaldava eksodentsuse iseloomulik tunnusjoon on sporeliininine olemasolu - kõrge molekulühend, millel on erakordne resistentsus; Kutinile füüsikalistele ja keemilistele omadustele. Madalamad taimed moodustavad zyospores ja aplanery (maitseta maitseid). Neil ei ole polüsahhariidi kest. EXODEE erineb hädaolukorra tugevusest, veekindlast, vastupidavust kõrgetele temperatuuridele, kemikaalidele ja mikroorganismidele. Seetõttu võivad maapealsete taimede vaidlused jääda elujõuliseks (mõnikord aastakümneid). See aitab kaasa idanevuse ebasoodsate perioodide kogemustele, mis on eriti oluline maapealse elupaigas. Tänu mikroskoopilistele suurustele talutavad nad sageli märkimisväärseid vahemaid. Seega vajavad elutingimustes elutingimustes vajalikke adaptiivseid muudatusi vaidluste struktuuri muutusi. Vaidluse abiga teostatakse liigid ja ebasoodsate tingimuste üleandmine. Shelli argument (eriti selle väliskihid) on ka geoloogilistes kujundustes hästi säilinud fossiilses olekus. Paljud pikaajalised taimed on teada ainult nende käskude jäänused.
Rohkem primitiivseid kõrgemaid taimi toodavad identseid vaidlusi (isospa) nii suuruse kui ka füsioloogiliste omadustena (morfoloogiliselt võrdsed voolutaimed). ISOSPort "idanema" jagamise soodsates tingimustes moodustuvad moodustumised (gametofüütide). Playans, Mängijad, sõnajalad ja PPI nad elavad iseseisvalt, sõltumata sporophyte. Toit erinevalt: autotroofne, (maapind, roheline), mükotroofne - sümbioos seente (maa all, värvitu) ja segatroofne (poolliin) Ingrowth.
Kuid maa-taimede hulgas on palju morfoloogiliselt eraldatud vorme, mis moodustasid vaidlused erinevad suurused ja alati funktsionaalsed omadused (heteropsosfer). Väikesed vaidlused (mikroseemport) idanemise ajal tekitavad meeste intensiivseid, suured (megasy) - naine. Need moodustavad vastavalt ühel või erinevatel isikutel vastavalt mikro- ja megaloprananate.
Kaasaegsete hobuste jaoks iseloomustavad füsioloogilised erinevused. Samad vaidlused erinevate troofiliste ja veevarustuse substraatide moodustavad erinevaid ülekasvatusi - meeste (halva kasvu tingimused), naised ja taglase (soodsate). See on tingitud asjaolust, et munaplaat koguneb embrüo jaoks vajalike toitainete pärast viljastamist. Füsioloogiline erinevus kaasaegsetes horsetailides on esivanemate morfoloogilise erinevuse "kaja".
Protsessis evolutsiooni, kõnniteel kaasneb vähendamine ülekasvanud, eriti meessoost. Sageli koosnevad nad ainult vegetatiivse keha ja ühe Aniardia ühest rakust. Naiste väljavool, lisaks munarakkude moodustamisele peaks tagama zygootide ohutuse ja embrüo arendamise ohutuse, mis varases staadiumis ei ole iseseisva majutuse võimeline. Post-asendis Plain-kujuline ja Fern-like Ingrowths arenevad sees Megapar või osaliselt väljaulatuv väljaspool kesta. Nad on usaldusväärsemalt kaitstud kui morfoloogiliselt samaväärsete taimede ülekasvamine. Plain-kujulise ja Fername megachers teostavad ka liikide lahendamise funktsiooni. Seemnete taimed MegaSpor ei jäta kunagi ema sporofüütist.
Maataimede puhul on põhiliselt uute mitmevärviliste reproduktiivorganite moodustamine või kasutute ja seksuaalsete protsesside organid (s.o Sporangiyev ja GameGangiev). See on tingitud vaidluse kaitsmisest ja välisest mõjust keerulise maapinna tingimustes. Väliste kihtide steriliseerimine esineb (moodustub seina); Ainult sisemine kangad on võimelised vaidluse ja mänge moodustama. Seina kindlalt hoiab niiskust ja kaitseb arendada vaidlusi ja geeks kuivatamisest välja. See funktsioon on üks tähtsamaid maapealse olemasolu all.
Sporali sein on ühekihiline või mitmekihiline. Tippide argumentide arv on erinev. Sporangia tonosfäärilistel kõrgematel taimedel moodustub vaid 8 vaidlust väga harva, tavaliselt vähemalt 32 vaidlust. Paljud on kaks korda või neljakordsed, mõned primitiivsed sõnajalad on kuni 15 000 vaidlust. Ühe positsioonide vormides moodustub igas mikrospoodis ka vähemalt 32 mikrosporte. Siiski moodustub üks megaspora megasprangias tavaliselt.
Kõrgemate taimede üldise taime organitel on alati kaks tüüpi: meeste - alternationalia ja naised - archegononia. Meeste seksuaalrakud moodustatakse Andifia - Spermatozoa ja Archegononia - naissoost sex rakud (naissoost gamets) - munarakk. Anterias on ovaalne või sferoidvorm. Aniardia ühekihilise seina all moodustub spermatogeenne kangas, mille flageli spermatosoid moodustuvad. Nende küpsemise ajaks ilmneb seina niiskuse juuresolekul ja spermatozoa lippude abil liikuda Archegraania suunas.
Arhegoonial on maitsev vorm. Ülemine kitsas osa on kaela, madalam laiendatud - kõht. Ühekihilise seina kaitse all kaela sees, emakakaela torukujulised rakud arenevad püksis - üks või kaks kõhu kanali rakku ja suurt sfäärilist muna. Munade valmimise ajaks söövad emakakaelad, kõhupiirkonnad, samuti seina ülemine rakk. Osa limaskesta paistab välja kaugemale archegonoonia piiridest. Lima sisaldab aineid, millel on positiivne kemotaktiline toime spermatozoa'le. Nad ujuvad kaela lima arhegooniumile ja liiguvad muna suunas.
Sporangia ja Gamentaania elutsükli jooksul piirduvad Gamentaania oma erinevate arengufaasidega (vormid või põlvkonnad). Samuti vähendab Gamenhai kõnniteel vormidest. Alsidia, Archegonia - Gtem, Velvichia on ennustanud. Kõik kaetud ääremeetmed ei ole üldse moodustunud.
Kõrgemad taimed ei moodusta isooamite ja heterogamiinide (erinevate põrandate seksuaalsete seksuaalsete rakkude erineva suurusega), leitud madalamates taimedes. Väli diferentseerimine kõrgemate taimede intensiivistunud ja viinud mängude terava dimorfismi. Munad kogunevad toitainete ja seega suuremate ja fikseeritud. Sperma ja sperma ei ole peaaegu vaba toitained. Seega on kõrgemaid taimi iseloomustavad anisogamets erinevad suuruse, liikuvuse aste.
Dumorfism mängudel on suur bioloogiline tähtsus. Meeste ja naissoost väravate vaheliste toitainete ebaühtlane jaotus annab suurema hulga ühendab võitlejad kui ühtlase jaotusega sama massi vahel toitainete vahel. Taimede arengu protsessis intensiivitati soojuse dimorfismi.
Seksuaalse diferentseerimise tugevdamisega suurenes meeste kaalude arv GAMENTAs ja naiste kaalude arv vastupidi vähenes. Archegonium pärines ühe suure ja fikseeritud munaga. Suur hulk Archegonia mune saab olla ainult anomaalsetel juhtudel. Spermatozoa suurenenud suurenemine suurendas seksuaalse protsessi tõenäosust. Sellega kaasnes nende suuruse vähenemine, mis aitas kaasa spermatosozoa liikumisele vee õhematesse kiledesse. Toitainete kontsentratsioon ühes suurtes munades aitab kaasa täieliku järglaste arengule.
Maataimede jaoks iseloomustab oogami. Väetamise protsess (Syngamy) esineb arhegonoonias. Primitiivsed kõrgemad taimed viljastamisel toimub toores ilm, vihma või rikkaliku kaste ajal. Liikuv meeste väravad leekide (spermatozoa) liikuda oma vee filmi ja REACH archeganiyev. Enamik hääletatud ja kõik kaetud taimed, tulemusena kohanemise elu tingimustes Sushi, seal oli eriline Oogami - Siphonogamy. Mees Gasets kaotavad liikuvuse. Väetamise protsess toimub ilma tilguti vedeliku söötmeta. Meeste kaalude kohaletoimetamise ja kaitse funktsiooni teostab õietolmutoru moodustumise tõttu hammetofüüdi (õietolmuterain). Need saavutavad suurepärase usaldusväärsuse viljastamise protsessi.
Seksuaalse protsessi tulemusena moodustub Zygoota, mis tekitab kõrgemate taimede multikulli idu. Nagu ZyGote, kõik tuumarakud iseloomustavad diploidikomplekt kromosoomid, mis sisaldavad pärilikku materjali vanemate mitte-nominaalse geneetilise olemuse. See tagab rohkem heterogeensem järglaste ilmumise geneetilises suhtumisel vanemate geenide rekombinatsiooni tõttu. Soodsad tingimused luuakse loomuliku valiku jaoks. Selles on see, et seksuaalse protsessi bioloogiline roll on.
Embrüo on järgmise põlvkonna noor sporofüüt. Tulevikus moodustatakse järk-järgult täiskasvanud sporreofüüt. Kõrgemate taimede funktsioon - mitmeraskulaarse embrüo olemasolu - Saksa botaanist V. Zimmerman, Nõukogude - A. Takhtajian ja American - A. Kronkovista nimetamine Harryobionta geriorganismide nimetamiseks (embrüoptsioon).
Kõige kõrgematel taimedel kriitilistes varajastes etappides areneb noor embrüo (sporophyte) emane gametaani sees emade oracte'i kaitse all. Ema organismi kõrgeimad eosed on gametofüüdi ja seemnetaimed - sporefit ise (kuna gametofit on äärmiselt vähendatud).
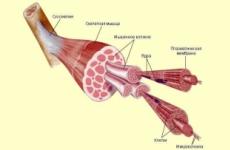
Kõrgemate taimede puhul on heteromorfne elutsükkel iseloomulik (joonis fig 1). Spore taimed, ta jätkab vaidlusi vaidlusi, seemne seemne seeme. Sisaldab kahte reproduktiivprotsessi: seksuaalne (sporris) ja seksuaalne. Kõrgemate taimede iseloomulik tunnusjoon on õige arengufaaside faaside õige muutmise olemasolu või vahelduva põlvkondade: seksuaalne (gametofüüt \u003d INFAPLOX \u003d haplite \u003d haplophazes) ja tiigel (sporofüüt \u003d diplofaat \u003d diplofaat). Nagu eespool märgitud, on need põlvkonnad paljudes kõrgemates taimedes (raamid, kummutid, sõnalased jne) mõned füsioloogiliselt iseseisvad organismid ja reproduktiivprotsessid jagatud mitte ainult ruumiliselt, vaid ka ajas.
Joonis fig. 1. Morfoloogiliselt ekvivalendi taime elutsükli diagramm - meessoost kujuga (lükopodiumiklambri) tavaline (Lycopodium Clavatum): 1 - Sporophyte (täiskasvanud taim); 2 - Spoorofülli sporangite; 3-6 - Ema-raku (spoori) vaidluse arendamine meioosi abil; 7 - Vaidlus; 8 - idanemine; 9 - Ozoeple Gametophyte (lüliti); 10 - Alteryidia; 11 - spermatozoa; 12 - Archegonium munarakkudega; 13 - Archegonium muna ja spermatozoomiga; 14 - Zygotesi lõhustumine (embrüo arendamine); 15 - ülekasvanud jäägid; 16 - Noor Sporophyte.
1-7,14,16 - elundid ja eoste struktuurid; 8-13.15 - Ingrowthi elundid ja struktuur.
MOV ja eriti seemnetetaimede puhul on üks põlvkondadest juhuslikult ja füsioloogilistes tingimustes, see vähendatakse tema keha. Glows domineerib gametofüüdi, kõik teised kõrgemad taimed - sporophyte, saavutades sageli suuri suurusi. Kõigis kõrgemates taimedes on vähendamise jagunemise koht (Meeo) vaidluse moodustamisel. Ühest diploidist emarakkudest moodustub vaidlus haploidide vaidluse tetrade (4-meioriori) tetrade poolt.
Safrofit lõpetab atogeenseerumise arengut mitmesuguste spooride moodustamisega vaidlustega. Kõige kõrgemate taimede puhul tekib spetsiaalsetes organites (sporangiye vedajad) välja arvatud sambla, välja arvatud sammal. Sporangiofonide päritolu, struktuur ja vorm on mitmekesised. Sagedamini on neil tasane lehtvorm, neid nimetatakse sporeofüllideks.
Pärast idanemist Haploidse vaidluse haploid seksuaalse põlvkonna moodustub - gametofit. Ainult vaidlusest pärit mugh-lonis arendab Protoonmea (esiplaanil) - kena või lamelli moodustumine, mis annab hametofüüdi alguse. Tsüklit korratakse.
Erinevalt madalamatest taimedest ei tee kõrgemate taimede vaidlused paljundusprotsessi ("enda paljundamine nagu"). Isegi juhtudel, kus vaidlused (morfoloogiliselt ekvivalentseerivates) ja Megaparis (morfoloogiliselt parallers) tehastes esinevad taimede ümberasustamise funktsiooni või naissoost gametofüütide (röhade) põlvkondade funktsiooni, mis nende morfoloogilistes, tsütoloogilistes ja funktsionaalses omadustes erinevad sporofitist järsult , sporangies, milles see moodustati.
Unastage kõrgemad taimed ja seksuaalse paljunemise. Zygoot ei arenda isikut sarnaselt vanemaga ja järgmise põlvkonna heteromorfse elutsükli - sporofüüdi.
Liikide uute omaduste tekkimine toimub ainult kahe reproduktiivprotsesside kombinatsiooni tulemusena - spooride ja seksuaalse protsessi kombinatsiooni tulemusena. Kõigekõrgemate vaidlustetaimede puhul viiakse reproduktsioon läbi gameto-spordialade tulemusena (kui domineerivad gametofüüdi - mugh-sarnase) või spoori-gamia (kui eosed domineerivad - ülejäänud vaidlused), (Sautkina, Polyksgenitova , 2001). Kõrgeimate spoori taimede, spooride ja seksuaalse protsessi demonteeritud ruumiliselt ja jätkata erinevate põlvkondade.
Seemnetaimede arengu protsessis esineb suur aromorfoos - megasoprangiy (megasinangius) spetsiaalse tüübiga - ilmub seemned (seemnete dip). Megaspi moodustamise protsessid, naissoost mänguofüüt, kaalu (munarakud), viljastamise protsessi, embrüo arendamist ühes orelis. Selle tulemusena muutub seemepea ise seemneks. Sofoori ja seksuaalse protsessi ei ole enam eraldatavad ja nende põhjal on eriline reprodutseerimisviis - seemne. Kaetud taimedes kombineeritakse need protsessid mitte ainult ruumiliselt, vaid ajas. Mõlemad protsesse järgneb üksteisele väga kiiresti, peaaegu katkematult. Ratsights on tugevalt vähenenud, nende arengu perioodid vähenevad järsult. Kogunenud taimed demonstreerivad eredalt elutsükli kestuse erinevuse lõpptulemust. Megaspora kaotas ka võime liigi lahendada, see funktsioon teostab seeme.
Enamik kõrgemaid taimi on tüüpiline taimse reprodutseerimisele. Erandiks on paljud lisatud ja kaetud-pruun-aastaste ja kaheaastaste taimede poolt. Loodusliku vegetatiivse reprodutseerimise vormid on äärmiselt mitmekesised ja sageli spetsialiseerunud, eriti kaetud taimedes. Kõrgemad taimed võivad korrutada vegetatiivsete elundite (Taltside, juurte, võrsete) ja nende osade abil. Mitte-spetsialiseerimata vegetatiivse reprodutseerimise vorm on laialt levinud killustumine juhuslike mehaaniliste tegurite mõju tagajärjel (tuule, loomade voogude, liikumise ja söömise mõju jne) ja selle tulemusena dieedi osa rakud. Spetsiaalsed vegetatiivsed reprodutseerimise vormid on ka haploidide ja diploidide põlvkondade jaoks laialdaselt oma domineerimisega. Spore taimed moodustavad neerude neerud, oksad, lehed, brood tales, juurdumine võrsed, adventtiivsed (nähtavad) neerud (ferns), maa-alused risoomid ja muud seadmed. Kaetud taimede lasteorganismide arenevad alati neerudest, mis on varustatud vegetatiivsete elundite erinevates osades (juured, varred, lehed). Nende lukustamine on sageli seotud organi mehaaniliste kahjustustega (looduslik ja kunstlik). Laialdaselt tavalised spetsialiseerunud organiseeritud organiseeritud organiseeritud organiseeritud õitsemise taimede reprodutseerimisel on risoomid, maa-alused ja õhuliinid (vuntsid), sibulad, tubekovitsa, mugulad, juurekoonused ja teised.
Kõrgemad taimed esinesid vetikatest, sest taimemaailma geoloogilises ajaloos eelnes kõrgemate taimede ajastu vetikate ajastule. Need hüpoteesid ilmusid X1X sajandi lõpus (F. Bower, F. Freach, R. VettsHTEIN). Esimesed usaldusväärsed maapealsed taimed on teada ainult vaidlustel ja on dateeritud Siluri perioodi algusesse paleosoilise ajastu (430 miljonit aastat tagasi). Konserveeritud mikro-löögi või trükkide järgi kirjeldati maapealseid taimi suurte ja madalamate Devoni setete hulgast. Esimesed kõrgemad taimed ühendatakse rühma rinofiit. Hoolimata struktuuri morfoloogilisest ja anatoomilisest lihtsusest olid nad tüüpilised maapealsed taimed. Neil oli kustetud epidermum koos lugude, arenenud juhtiva süsteemiga, mitmesugused spoorid tugeva kaitsekoorega. Seetõttu võib eeldada, et kuivatusprotsess algas oluliselt varem - varases paleosouses (Ordovikis või Kambriumi perioodides).
Kõrgematel taimedel on terviklik funktsioonikompleks, mis viitab nende päritolu ühtsusele ühest esivanematest rühmast (monofiileetiline päritolu). Siiski on palju seisukohti seisukohti polüfileetiline päritolu, sealhulgas raames üksikute rühmade kõrgemate taimede (mugh-kujuline, kaetud). Nende seisukohtade kohaselt valdas vetikate allika rühm erinevaid taasesitussüklit ja andsid kaks sõltumatut arengut.
Pikka aega peeti kõrgemate taimede esialgse rühmana pruuni vetikate (varre, potonier). Nende seisukohtade toetaja oli ka K.I. Meyer. Neid iseloomustavad keerulised arengutsüklid, sealhulgas heteromorfsed. On igasuguseid põlvkonna vaheldumisi. See on üks keerulisemaid vetikate rühmade, nii väliselt kui ka sisemiselt. Seda iseloomustab multikulaarne üks, mis on sageli raske hautatud ja leasareorganite kõrgel tasemel raske. Paljudel pruuntel vetikal on koe struktuur (assimilatiivsed, päringud, mehaanilised ja juhtivad kuded eristatakse). Selle osakonna esindajad kohtuvad mitmevärviliste spataansete ja Gamentangiaga. Kuid looduses on seadusega "mitte-spetsialiseerimata esivanem" sageli kehtiv. Lisaks iseloomustas esimesi kõrgemaid taimi primitiivse välise ja siseruumi struktuuriga. Üks raskusi selle hüpoteeside vastuvõtmiseks on pigmendi kompositsiooni ja vabade toitainete erinevused: pruunid vetikal on klorofülli a ja c (viimane ei ole tuvastatud), lisamise pigment fukoksantiin, aktsiatooted on lamineriin ja kuue- Katke alkoholi (neil ei ole tärklist). Brown vetikad on ka üksnes meri, mitte mageveeorganismide merel. Arhegonaalsete taimede hulgas ei ole mere taimestiku esindajaid. Ainult seas kaetud soolalahusega 20-30 liiki elavad soolatud vetes. Kahtlemata on need sekundaarsed kõrge organiseeritud taimed.
Kahekümnenda sajandi teisel poolel toimunud uue teabe tulekuga hakkas roheline vetikad kaaluma uuesti kõrgemate taimede esivanematena (L. Stebbins; M. Shafedo jne). Kõrgemate taimede ja roheliste vetikate puhul iseloomustab sarnaste pigmentide olemasolu sarnaste pigmentide olemasolu (peamine fotosünteetiline pigment on klorofülli A, a-klorofülli a-ja β-karoteenina, sarnased ksentofüllid), nende plast Hästi väljendunud süsteem sisemiste membraanide. Assimilatsiooni saadus (peamine vaba süsivesikute) on tärklis, mis lükati edasi kloroplastides, mitte tsütoplasmas, nagu teised fotosünteetilised eukarüootid. Kõrgemate taimede ja mõne rohelise vetikate puhul on rakuseina kõige olulisem komponent tselluloos. Samuti esineb mõningate rinofiidi sarnasust mõne rohelise vetikaga filiaali olemuse järgi. Eraldi kaasaegsed hetofoorid (tellida katofoorel) on multi-kammer-gamente. Mõnede esindajate elutsüklite sarnasus on ka sarnasus. Koos liikuvate zoosporalitega on roheliste vetikate fikseeritud aplanery, nii kõrgemate taimede iseloomulik. Nad elavad peamiselt mageveehooldustes ja maal. Morfoloogiline ja keskkonna mitmekesisus, nende elustrateegiate mitmekesisus võimaldas neil areneda erinevates suundades. Niisiis, enamiku kaasaegse botaanika sõnul võivad kõrgemate taimede tõenäolised esivanemad olla magevee- või solanovaansed multikulaarsed rohelised vetikad heterootikaga (häbistatud) Taloma struktuuriga.
Hüpotees päritolu kõrgemate taimede päritolu organismidest sarnanevad elava Haroviga ka laenud laialt levinud. Mõlemad rühmad koondavad rakkude pektiiniplaadi moodustumise olemust (fragmoplat on kaasatud - mikrotuubulisüsteem, mis asub mitootilise spindli ekraatoritasandil). Samad arengusuunad - keskusest perifeeriale (tsentrifugaal). Sarnaselt intertsellulaarse plaadi moodustumise tüübi leidub ka mõningate ULotrics'i klassi esindajate hulgas, mida iseloomustab ökoloogiline struktuur Taloma (NITAL, vähem sagedamini plaat või torukujuline).
Koos keerulise morfoloogilise likvideerimisega Taloma struktuuri, harufits iseloomustab multiklular organite seksuaalse paljunemise, samuti kõrgemate taimede. Nende olulised iseärasused on ka prootoni etapi olemasolu või auaste, arenguprotsessis, tala haploidne iseloom (züootiline vähendamine). Nad elavad veekogudes värske ja soolase veega veega. Mõned piirduvad maapealsete niiskete elupaikadega. Fossiilsete matkide vetikate jäägid on tuntud paleosoilise ajastu silurperioodi setetest. Samuti eeldatakse, et nad oleksid võinud juhtuda mõnede kõrgelt organiseeritud vastastikusetest rohelistest vetikatest, mis sarnanevad kaasaegsete hetofooridega.
Muud pilgud tõlgitakse kirjandusse kõrgemate taimede päritolu. Arvatakse, et seal võib olla mingi hüpoteetiline rühm on esivanemad, ühendades märke pruuni ja rohelise vetikate. Teised vähem levinud hüpoteesid on teada.
Samuti soovitati, et üleminek kõrgemate taimede esivanemate üleminek maapinnapõhise olemasolu tingimustes aitas oluliselt kaasa seened sümbioosile (sümbogeneesi teooria). Nagu te teate, on looduses laialt levinud sümbioos seentega. Intratsellulaarse taseme juures maa-aluste asutustega (endomicoroos) on kõige kõrgemate taimede jaoks iseloomulik. Paleosoilise ajastu alguses oli raske korraldada mitmevärviline vetikad hakkas rannikumaad asuma endomükorisatsiooniliidu koostises. Vanimate kõrgemate taimede jääkide uuringud näitasid, et ETDOMICRIZA kohtus mitte vähem kui kaasaegses. GIF-seente olemasolu maa-alustes orelistes kudedes aitas ilmselt kaasa mineraalsete ainete intensiivsemat kasutamist, eriti vaeste toitainetega suletud fosfaate, alustasid substraadid paleosoilise ajastu. Lisaks eeldatakse, et see võib pakkuda ka parima vee imendumist, suurendada kõrgeima taime jätkusuutlikkust kuivatamisse, mis on elu tingimustes äärmiselt oluline. Sarnaseid suhteid võib täheldada kaasaegsete taimede näitel, esimene, kes asuvad äärmiselt halva pinnase. Sellistel tingimustel on sellistes tingimustes palju rohkem võimalusi ellujäämise võimalusi. Seega on võimalik, et esimene lahendati maale mitte üks organism, vaid terve süsiootiline kompleks.
Eeltingimused maa taimede välimuseks olid mitmed. Teise maailma sõltumatu kursus valmistas ette uute ja täiustatud vormide tekkimise. Sea vetikate fotosünteesi tulemusena suurenes hapnikusisaldus, mis võimaldas arendada elu maa. Proterozoic Eru (900 miljonit aastat tagasi) hapniku kontsentratsioon atmosfääris oli ainult 0,001 kaasaegse taseme, Kambriumi (esimene periood paleosoeerumite ajastu) - 0,01 ja hõbedas - juba 0,1. Hapnikusisalduse suurenemine korrelatsioonis osoonikihi moodustamiseks, mis säilitab mõne ultraviolettkiirguse. Esialgsetes etappides arengu elu maa peal, see aitas kaasa kujunemise bioloogiliste makromolekulide ja samal ajal tegi teguriks piirates evolutsiooni puudumisel piisava hapniku atmosfääris. See on peamiselt vajalik kerneli ja rakkude jagamiseks.
Maapealsete taimede tekkimine langeb kokku fenoolühendite metabolismi, kaasa arvatud tubüül-ainete, antrotsüaniinide, flavonoidide jne väljatöötamisega. Nad reguleerivad kasvuprotsesse, aitama kaasa taimede kaitsereaktsioonide väljatöötamisele, sealhulgas mutageensetest teguritest - ioniseerivast kiirgusest , ultraviolettkiirgus, mõned keemilised ained.
Hiljuti omandavad üha rohkem toetajaid hüpoteesi, et madalama sorte on õppinud madalamaid organisme - erinevaid vetikad, seente ja baktereid, moodustades maapealse ökosüsteeme koos. Nad valmistasid ette substraadi, õppis hiljem taimede poolt. Palekomeerid on juba varajasest prepambriast teada.
FOSSIL FLORA S.V kuulsate ummikute kohaselt. Meyena (1987) on tõenäolisem, et kõrgemate taimede moodustamise protsess ei juhtunud vetikate väljumise ajal maale, vaid Sushi vetikate elanikkonnast. Nende moodustamise aeg tuleks seostada ränidioksiidi eelmiste perioodidega.
Paleosoilise ajastu alguses toimusid ulatuslikele territooriumidele suured moodustumisprotsessid. Skandinaavia mäed, Sayans, Altai, Põhja-Tian-Shan jne. See põhjustas paljude merede järkjärgulist kooriku ja Sushi välimust endiste madalate veehoidlate kohas. Kuna mered ületavad, olid madalas vees asunud madalas vees arveldatud vetikad maale. Ellu jäänud ainult need taimed, kes suutsid kohaneda uute elutingimustega. Kõrgemate taimede esivanemad olid vajalikud kõigepealt kohaneda eluga haarajas, siis magevees, suudmes, madalas vees või reservuaaride märjalistel kaldal.
Uus elupaik oli põhimõtteliselt erinev esialgsest veest. See on iseloomulik: päikesekiirgus, niiskuse puudujääk, kahefaasilise pinnase-õhu kandja keeruka kontraste. Üks peamised hetked varajases staadiumis väljumise maad oli moodustamine vastupidav kaitsekaaned, kuna niiskuse puudujääk oli kõige olulisem kriitiline tegur arengu maapinna. Vaidlused olid võimelised levima sushi pinnale tuule poolt ja taluma kuivades tingimustes. Dispersiooni puhul on vaidlus vajalik, et sporangiiad tõstatatakse substraadi kohal. Seetõttu kaasnes sporofüüdi arendamine selle suuruse suurenemisega. See nõudis rohkem õhu ja mineraalse toitumise tooteid. Sellest tulenev taimse pinna suurenemine saavutati selle likvideerimisega kõige lihtsamal viisil - maapinna ja maa-aluste telgede Willy hargnemise. Taimede suuruse suurenemisega ilmusid struktuurid, mis aitavad kaasa vaidluse tõhusamale vabastamisele ja hajumisele. Teine olulised evolutsioonilised protsessid aitavad kaasa Sushi maandumisele Kutini taimede biosünteesi ja üldiselt keerulise spetsialiseeritud kanga moodustumist - eraldistega epidermis. See suudab kaitsta maataimede kuivatamist ja läbib gaasivahetust. Kõrgemad taimed stabiliseerus niiskusesisaldus ja muutus suhteliselt sõltumatu atmosfääris ja pinnas niiskuse kõikumisest. Madalamatel taimedel ei ole veevahetus stabiliseerunud. Nende oluliste protsesside intensiivsus sõltub täielikult elupaiga niiskuse olemasolust. Kui esinemise ajal põudas, kaotavad nad niiskuse ja satuvad anabiosisse.
Maapealsete taimede morfoloogiline lammutamine toimus roheliste vetikate heterootrichalilisest talus. Arvatakse, et nende teritamisosi andsid kihistumise alguse (Talombom) vormid ja kasvavad radiaalsed. Plaadi taltomas bioloogilises mõttes osutus vähendatuks, kuna need põhjustaksid süveneva konkurentsi valguse jaoks. Kasvavad alad, vastupidi, said täiendavaid arengut ja moodustasid veelgi radiaalseid aksiaalstruktuure.
Kuna kõrgemate taimede väljapääs maale töötati välja kahes peamistes suundades ja moodustasid kaks peamist evolutsiooni oksad: haploid ja diploid.
Kõrgemate taimede arengu Haploidse haru iseloomustab gametofüüdi progressiivne areng ja seda esindab Moss. Koos seksuaalse protsessi osutamisega täidab Gametofit vegetatiivsete organite peamised funktsioonid - fotosünteesi, veevarustuse ja mineraalse toitumise funktsioonid. Nad järk-järgult parandasid, muutusid keerulisemaks, suurendades assimilatsiooni pinda, morfoloogiliselt häiritud areneva sporofüüdi vaidluse pakkumise ja hariduse tagamiseks. Sporophyte vastupidi on läbinud vähendamise. See piirdus enamasti spoori vormiga ja see ei ole iseseisvalt elav tootmine. Parema dispersiooni jaoks töötati välja vaidlus erinevate seadmete poolt.
Seksuaalne protsess sammal toimub tilguti vedeliku niiskuse juuresolekul (meeste mobiiltelefonid - Bow-isase spermatozoa) juuresolekul. Seetõttu on gametofit sageli seotud niiskete elupaikadega ja ei suuda saavutada suuri suurusi. Haploid-gametofüütil on ka väiksem geneetiline potentsiaal kui diploidne sprofüüt. Seetõttu on kõrgemate taimede arengu rida gamenaofüüdi domineerimisega külg, surnud lõpuni.
Kõik muud taasesituse tsükli kõrgemad taimed domineerib sporofüüdi. Diploidsed kromosoomide komplekt koos assimilatsiooni aktiveerimisega on laiendanud moodustumisprotsesside võimalusi. Sporophyte maatingimustes osutus palju elujõulisemaks.
Maapealsete taimede arengu pöördepunktiks oli rakkude ilmnemine ligniini sünteesiks. Nad moodustasid juhtivad ja toetavad kuded. Alates Devoni perioodil muutus sporofüüdi maa-alused osad imendumise ja konsolideerimise funktsioonide funktsioonide täitmiseks. Kõrgemate taimede ülalmainitud osade kohta moodustati lehed. Niisiis, kuna kõrgemate taimede keha suurus suureneb anatoomiline ja morfoloogiline diferentseerimine keeruliste spetsialiseeritud kudede ja elundite moodustumiseni. See tugevdas kõrgemate taimede positsiooni maapealsetes keskmistes ja aitas kaasa tõhusama fotosünteesi. Rikkalik hargnemine ja suurte sporofüüdi suuruste loomine on korduvalt põhjustanud diasporaa tohutut tootlikkust ja nende tõhusat lahendamist. Enamikus taimekogukondades domineerivad sporofüüdid.
Gametofit, vastupidi, arengu ajal vähenes järk-järgult ja lihtsustatud. TalnoSpectoris lennukite, Mängijad, Ferns Gametophytes on kujul väikese diferentseerumata või nõrgalt diferentseeritud roheline tilla, mis on saanud nimega kattuva või protalliumi (protellium). Gamemarketofyte'i maksimaalne vähendamine on seotud korruste eraldamisega. Ülesanne piirdub peamise funktsiooni rakendamisega - seksuaalse protsessi tagamine. Mõlemad tüüpi sama seksuaalsete gametofüütide (meessoost ja naine) on palju väiksemad kui gametofüütide evivori maapealsete taimede. Paratingi ajal arendavad nad vaidlusi kesta all ja rullis väljaspool seda.
Sama seksuaalsete gametofüütide vähenemine ja lihtsustamine toimus kiirendatud tempo arengu protsessis. Nad kaotasid klorofülli ja nende areng oli tingitud sporofüüdi toitainetest. Gamenaofüüdi suurimat vähendamist täheldatakse seemnetaimedel. Mees Gametofüütide esindab õietolm teravilja, naissoost gametofyte Vicred - primaarne haploid endosperm, kaetud-äärega - embrüonaalne kott seemikud.
Sushi arendamise esimeses etapis maapealsete taimede poolt esines konkurentsi. Geoloogiliste setete argumendi monotoonsus ja arvukus näitab nende tähtsusetu mitmekesisust ja sushi kiiret omandamist. Devoni ajal (400-345 miljonit aastat tagasi) muutusid evolutsiooni diploidiliini kõrgeimad spoori taimed arvukamaks ja mitmekesisemaks. Esimest korda on morfoloogiline kõnniteel. Tulevikus tekib see korduvalt erinevates veressioonitehastes. Ühekordne vastupidavus on oluline bioloogiline väärtus. Kõrgemates organiseeritud taimedes ainult heterosepooria põhjal on õded, seemned jne.
Taimede vormide järsk diferentseerimine toimus keskel. Soodsam rhinofiit asendatakse puu-nagu kõrge (kuni 40 m kõrgusega) koos tavaliste ja pehmendaja kujuga. Hilise Devoni aja jooksul moodustavad puutaimed reaalsed metsad. Suhteliselt lühikese ajavahemiku jooksul (hilja Devon - süsinik (söe periood)) ilmuvad paljude taksonoomiliste fernatsioonirühmade esindajad. Nad hakkasid domineerima elupaiga sobiva sushi osa. Planet hakkas roheliselt. Seda perioodi nimetatakse õigustatult Ferni ajaks. Kõige suuremad arenduskaadrid jõudsid söe perioodi lõpus. Kõrge puu-sarnased plano-sarnased (Lepidodendrons, Sigillaria jne), Mäjad (Calamiidid), Fern ja seemnevärvid moodustasid lopsakas taimekogukonnad, paljudes aspektides sarnanete kaasaegsete troopika taimede maailmaga. Sel ajal ilmus naughty.
Paleosoeerumise ajastu lõpus (Permi periood) hakkas peaaegu kõikjal maa peal valitsema. Nad olid varustatud neile, kes valitsevad Fern. Mesozoicis (hääletatud ajastu) on suurim mitmekesiruumide suurim valik. Arvatakse, et taimede järsk muutus on suures osas seotud kliimakuivuse suurendamisega. Arenenud juhtiva süsteemiga kaasnevam on kohandatud muutunud elupaikade tingimustele. Olulisi kohanemisfunktsioone iseloomustab sisemise väetamise protsess õietolmutoru abil, mis on iseloomulik paljude esindajatega. Ja lõpuks, neil on olnud vähem ja seemneid, mis toidavad sporofüüdi idu ja kaitsevad selle maaelu muutuste eest. Need on seemnetaimede peamised bioloogilised eelised.
Veelgi kohandatud keskkonnatingimustele on õitsemise või kaetud taimed. See lööb nende suuruse, eluvormide, tolmeldamise seadmete mitmekesisuse mitmekesisust, diasporaate jaotus, kõrvaltoimete ülekandmiseks jne. Kõigil nendel funktsioonidel on lubatud täielikult realiseerida nende evolutsioonilise kohanemispotentsiaali. Nad osutusid ainsaks osaks taimedeks, mis suudavad moodustada keerulisi mitmetasandikogukondi (eriti troopika kaetud metsades), mis koosnevad peamiselt ja mõnikord nende esindajatest peaaegu täielikult. See aitas kaasa elupaikade intensiivsemale ja täielikumale kasutamisele ning edukamatele territooriumide vallutamisele. Ükski taimede rühm ei suutnud selliseid erinevaid seadmeid teatud keskkonnateguritega arendada. Ainult nad suutsid merel keskkonda viivitamatult kapten - merekeste soolavees, kümneid kaetud sildade esindajad kasvavad koos vetikatega. Mesozoic ajastu kriidiperioodi alguses ilmuvad (neid hinnatakse nende fossiilsete jääkide poolt usaldusväärselt kindlaksmääratud), kui kriidi lõpuks domineerivad nad tavaliselt enamikus ökosüsteemidest. Suhteliselt lühikese aja jooksul geoloogilise aja jooksul, arvutatud kümne kahe miljoni aasta jooksul, nad läbisid peamise evolutsioonilise diferentseerimise ja laialdaselt levinud kogu maailmas, kiiresti jõudnud Arctic ja Antarktika. Nende domineerimine sushi taimekaitses jätkuvalt praegu. Biosfääri metaboolsete protsesside ja energia ümberkujundamise käigus sõltuvad atmosfääri, kliima, sushi veerežiimi gaasikoostis, mullaprotsesside olemus sõltub nende elatist. Enamik maapealseid loomi eksisteerivad ainult kaetud silla tõttu. Nad moodustavad nende elupaikade, seostatakse nendega erinevaid troofilisi ja muid nõustamisühendusi. Paljud loomsed rühmad tekkisid ainult siis, kui õitsemine hakkas maal domineerima. Paljude lülijalgsete (eriti putukate) ja mõnede selgroogsete (eriti lindude) jaoks on kaetud sildadega seotud evolutsioon iseloomulik. Inimene kui bioloogiline seisukoht suutis tekkida ja eksisteerida ainult kaetud silla arvukuse tõttu.
Nagu taimestiku maailmas, siis paralleelse arengusuunda täheldatakse ka loomade maailmas - alates muna kuuluvast Viviors. Taimed - Story-vormingust seemnete moodustamiseni. Seemnete taimed ületavad vaidluse. Lõppude lõpuks, tohutu enamus vaidlust, ei leidnud soodsaid tingimusi, sureb. Sporitel ei ole piisavalt toitaineid. Gametofüüdi arendamine, vaidluste väetamise protsess nõuavad ka teatavaid tingimusi, mida maal on alati võimalik pakkuda. Seemned on tavaliselt toitainete tarnimine (nende primaarse haploidse endospermi, kuuma silla - sekundaarse triigeeri endospermi, peispermi või väga idanemise iseenesest). Seemnekoori, mis on sagedamini integraalsete kudede (Intericuments) kudedest, on usaldusväärne kaitse keskmise kahjulike mõjude eest. Seemned on palju kohandatud kui vaidlus (üks rakk). Enamiku seemnete puhul on iseloomulik enam-vähem pikk puhkeaeg. Ülejäänud perioodil on palju bioloogilist tähtsust, sest see võimaldab kogeda aasta ebasoodsat aega ja aitab kaasa ka kaugemale lahendusele. Seemned - kõige kohandatud elundite taim lahendamiseks.
Vahepeal vähenes gamestofitide genereerimine järk-järgult suurusega ja muutus üha sõltuvaks toitumise ja kaitse spooridest.
Seega tähistati maapealse või kõrgemate taimede välimus uue ajastu alguses planeedi elus. Sushi taimede arendamisel kaasnes uute loomaliikide tekkimine. Taimede ja loomade konjugatiivne areng tõi kaasa eluvormide tohutu mitmekesisuse maa peal, muutis oma välimust. Kõrgemaid taimi kasutatakse ka talus ja igapäevaelus laialdaselt. Pealegi on peaaegu kõik kultuurijaamad, välja arvatud üksikute punase (porfyra \u200b\u200bjne) ja roheliste vetikate (kloralla, scentestsmus jne) erandiga kõrgemad taimed.
Perekonna kõrgemad taimed ühendab vähemalt 350 tuhat liiki. Enamik süstemaatika on jagatud 8 osakondadeks: mugh-kujuline, rimey-kujuline, tavaline, horsetails, fern, psülliline, hääletus, kaetud või õistaimed. Kuid kaasaegsetes akadeemilistes väljaannetes varieerub osakondade hulk 5 kuni 14. Varjab fossiilsete taimede, paleobotaanistide (SV Meyen et al.) Tuleb märkida, et seal ei ole ühtegi sündi, mis hõivata vahepealse asendi vahel tavalise kujuga ja fernation, kombineerige erinevate osakondade märke, tuues lähemale. Seetõttu on need sageli ühes osakonnas. Lisaks on nende taasesituse tsükli spetsiifilised spetsiifilised spetsiifilised spetsiifilised ained sõltumatult elavate heteromorfsete põlvkondade järjepidev vaheldumine. Ronition-on vastupidi, sageli eraldatud 2 või 3 sõltumatu osakonnad, moughen-sarnased - 3, 4 osakonnad. See kehtib ka hääletatud, jagatuna 5 osakonda (A.l. Takhtajian). Hiljuti tõsteti hiljuti kaks vanimaid külmutaimed esile - Archaeopteridofyta ja arheopteridofyta ja arheoSpermatophyta (N.S. Snigrevskaya).
Laevade ja (või) sõnul nimetatakse kõigi osakondade esindajaid, välja arvatud sambla, nimetatakse sageli kõrgemaid veresoontetaimi (tracheophyta). Ei ole vastastikku arenenud sisemist juhtivat süsteemi. Seemnetaimedele viitavad ja kaetud sild, ülejäänud osakonnad on kõrgeima vaidluse. Lisaks pilkadele ja seemnetele ühendatakse ülejäänud osakondade esindajad mõnikord vaskulaarsete taimede vaidluste rühma. Tänu vähendamise archegraniyev, kaetud sillad on vaidlustatud kõigi teiste osakondade - archegranoletion taimed.
Valgevene kõrgemate taimede determinantides "näidatud taimede elupaikade analüüs näitas, et Flora Valgevene umbes 1220 kohaliku või aborigeense veresoonte taimed. Selle tulemusena rikkumise terviklikkuse taimkatte tõttu aktiivse majandustegevuse isiku, loomulik taimestik on ammendumine ja samal ajal selle rikastamise tõttu hääletuse või adventti liikide. Mõned liigid esitatakse juhuslikult, eriti transpordiga (raudtee-, autotööstus, vesi, lennundus). Umbes kolmandik Valgevenest on hõivatud kultiveeritud taimed (toit, sööt, ravim, dekoratiivne, tehniline). Paljud neist tungivad sünaantoopilised elupaigad (jäätmete, prügilate, rauast, maanteede ja põlluteed jne). D. I. Tretyakova teadusuuringute tulemuste kohaselt sisaldab taimestikuõpetaja Flora osa rohkem kui 800 liigi. Lisaks veresoonte taimedele Blara Valgevene umbes 430 liiki sambla (G.f. Rykovsky, O.M. Maslovsky). Seega on praegu Valgevene Flora üle 2450 kõrgemate taimede liiki.
Looduslike ressursside intensiivse kasutamise tingimustes on taimemaailma kaitse probleem üha olulisemaks. See on keskkonnakaitse lahutamatu osa. See on terviklik ratsionaalse kasutamise meetmete süsteem, loodusressursside taastamine ja korrutamine. Ettevõtte arengu praegusel etapil on inimese majandustegevus üks loodusaressursside olukorra määravaid tegureid. Teadusliku meetmete süsteemi loomine, mis vastab looduse ja ühiskonna arengu põhiõigustele ning nende kasutuselevõtule ja looduskaitse objektiks. Punane raamat on peamine dokument, mille kohaselt viiakse läbi haruldaste ja ohustatud liikide õiguskaitse. Valgevene punases raamatus oli loetletud 171 kõrgema taimeliiki (sh 15 - moughen-kujuline, 12 - spoori vaskulaarne ja 144 - seemned: 1 vaade - hääletatud, 143 - kaetud). Alates 2000. aastast toimub töö Valgevene punase raamatu kolmanda raamatu ettevalmistamisel, mis on kavas lisada 200 kõrgema vaskulaarse taimeliiki. Töö toimub Valgevene rohelise raamatu loomise kohta, mis määrab haruldaste ja kaduvate taimede kogukondade seisundi. 1. juunil 2004 Valgevene spetsiaalselt kaitstud looduslike territooriumide olemasolev süsteem sisaldab umbes 1500 rajatisi (sh 1 biosfääri reservi, 5 rahvusparki, vabariiklasi reservi ja kohalikku olulisust jne). Nad hõivavad umbes 8% Valgevene territooriumilt, mis on rahvusvaheliste soovituste optimaalse taseme lähedal.
Kõrgeimad taimed hõlmavad kõiki maapealse lehtede taimed, mis toovad vaidlused või seemned.
Kõrgemate ja madalamate taimede peamised erinevused:
1) elupaiga: Madalaim - vees, kõrgeim - enamasti maa.
2) mitmesuguste kudede kõrgemate taimede arendamine- juhtiv, mehaaniline, kate, millest asutused koosnevad.
3) vegetatiivsete elundite olemasolu kõrgemates taimedes:
- Juurduma - kinnitamine pinnase ja vee mineraalse toitumise
- Leht- fotosüntees
- Tüvi - transportida in-in-in (kasvavalt ja allapoole voolu)
(varred lehed + neerud \u003d põgeneda)
4) Kõrgematel taimedel on kattelapp - Epidermis esinevad kaitsefunktsioonid
5) kõrgema taime varre tugevdatud mehaaniline stabiilsus paksuse rakuseina tõttu immutatud ligniiniga.
6) Paljundusorganid: Enamik madalamaid taimi on üksiklikud, kõrgemates taimedes - multikulaarne. Kõrgemate taimede tervitusorganid moodustatakse erinevatel põlvkondadel: ON gametofit (Aniardia ja archegononia) ja edasi sporofüüt(Spoorgia).
Tuginedes märke, mis on saadaval kõrgemate taimede nimetatakse: eraldatud, idanemise, võrsed, pikk ja vaskulaarne taimed.
Vaskulaarsed taimed - kõik kõrgemad taimed, välja arvatud samblad.
Kõrgemad taimed toimusid rohelised, magevee- või sirged heterotrichny vetikad. Esimesed kõrgemad taimed olid rynofiti - Bellless, biokotoomilised taimed. Nende taimede piiratud sprigsi kutsuti telkes.
Arengutsükli kõigist kõrgemate taimede, välja arvatud mus, valitseb sporophyte. Ainult Mukhis valitseb Gametophyte sprofüüdi üle.
Taimed on olemas : 1) Ravnoforovye - Nad moodustavad samu vaidlusi ja iga vaidlus idaneb mitmekesine gamemitofüüt.
2) üksikasjalikult- Naine gametofüüt on moodustatud naiste vaidlusest meestest meestest - meeste gametofitist.
Ühe tuuma, haploidse rakk (N) koos 2 kestaga.
Spore taimed:
Rhinofiit - Fossiilsed taimed (Rhyniophyta)
Massiline
Plenyophid
Playvoid
Herks
Fern-sarnane
Väemisel on vajalik vesi
Kõrgemad seemneseadmed:
Õitsemisosakond (kaetud soolalahus)
Väetamisega ei ole vaja vett
1. Kehaomaduste osakond Bryophyta osakond Bryophyta - Mossoid
Massiline - Kõige primitiivsem, vanim kõrgemate taimede rühm ilmus umbes 400 miljonit aastat tagasi.
Liikide arv: Praegu on brioloogid kirjeldanud umbes 20 tuhat liiki sambla liiki.
Võttes sammal:mosoidid jaotatakse kõikjal (settida pinnase, kivimite, kännude, puude), välja arvatud merede ja väga soolalahuse pinnase, leidub Antarktikas. Mossi eelistavad varjutatud niisutatud kohti.
Keha struktuur Makhov: Mossi - madalal vaimustunud mitmeaastased rohttaimed suurus 1 mm kuni mitu sentimeetrit, harvemini kuni 60 cm või rohkem. MOSS keha jagatakse kas varre (cauldia) ja väikesteks lehtedeks (fännus), näiteks Sfagnum ja Cucushkin vilgub või on esindatud kihiga, mis ei ole jagatud organiteks (märtsiks). Kõigi mossoidide iseloomulik märk - Rootsi ei ole. Vee imendumine ja substraadi kinnitus viiakse läbi nende risoomide kujuga, mis esindavad epidermis tekkimist. Vee imendumine ja aurustamine viiakse läbi kogu gametofüüdi kogu pinnaga.
Moch-like ei ole arenenud juhtivat süsteemi (tracheid, laevad, sõela torud). Seal on ühe magamistoaga ja kodumaiste taimede. Nende sisemine struktuur on suhteliselt lihtne. Mosoide, samuti kõigi kõrgemate taimede puhul on seksuaalsete ja võimsamate põlvkondade õige vaheldumine iseloomulik. Arengutsüklis domineerib haploidne gametofüüt (kujutab endast taime peamist keha). Sporophyte - ei sisalda klorofülli ja elukestvalt külge gamemarketofüüdi ja toidab kulu arvelt.
Moss'i areng on väga huvitav. Väetamine on võimalik ainult vee juuresolekul, sest spermatozoa võib selles liikuda. Meeste rakud leekidega moodustuvad ühel taimel, erinevates taimedes, kõige sagedamini, suured naissoost rakud küpsevad. Vihma või udu ajal, liikudes meeste rakkude tilk vett kiirustasid naiselik ja ühendada nendega. Väemalt naissoost rakkudest (zygotes) areneb sporofüüdi, mida nimetatakse sportlane (ta on kasti jalagalaiendatud allosas suu - GautoriaMis abi, mille ta uppumine Gamenaofyte, elab kulul see).
(Calipra-Satatake kõht archegonia)
Gametofüüdi ja sporofyte ühendus on väga piiratud. Gametophyte mitte ainult toidab, vaid ka kaitseb ka spoori genereerimist, aitab dispersioon vaidlus ("vale jalga" tõstatab kasti üle taime, archeganius, võttes maha oma kõht, katab kasti).
Kast moodustab tohutu arvu vaidlusi. Iga vaidlus on väiksem manna. Kui vaidlused valmivad, avab kast kaane või väikeste poorid moodustub selles, mille kaudu vaidlused lennata vabadusele. Üks kord soodsates tingimustes idaneb vaidlus. Moss'i individuaalne elu algab idanemise idanevusega. Kõige sagedamini, kusjuures turse, eksinikradade vaidluste lõhkemise ja panthin koos vaidluste sisuga koostatakse ja annab ühe rea lõime või ühekihilise plaadi kandmise alguse. Seda Gametofyt Arengu algstaadiumist nimetatakse protooniline (Kreekast. Protos - esmane, nesa - niit). See on kas järk-järgult pöörates täiskasvanud tllamic gametofüüdi (kariloomade) või neerude, tekitades täiskasvanud lehtede gametofüüdi protorgiable).
Vegetatiivselt, Moss korrutatakse eriorganite abiga (neerud neerud, lehed, lehed, osi, oksad), vegetatiivselt võimelised mitmekordistama ja sporofüüdi (jalg).
Mosssi suudab koguda palju aineid, sealhulgas radioaktiivseid aineid. Mõned Mossoids (Sphagnum) on antibiootikumide omadused ja neid kasutatakse meditsiinis. Turbamaksed moodustavad peamiselt Sphagnum Moss on juba ammu töötatud kui kütuse ja orgaaniliste väetiste allikana. Mosoidosakond on jagatud kolme klassi: 1) rozhkvesti (Antoker); 2 )Peches(Marsšus mitmekesine); 3) lehtede sammal(Cukushkin Luck, Sfagnum).
Orel on osa taimest, millel on teatud väline (morfoloogiline) ja sisemine (anatoomiline) struktuur vastavalt selle teostatavale funktsioonile. Taimede vegetatiivsed ja reproduktiivsed organid eristatakse.
Peamised vegetatiivsed organid on juured ja põgenemine (varred lehed). Nad pakuvad toiduprotsesse ja selles lahustunud aineid ja vegetatiivset reprodutseerimist.
Reproduktiivsed organid (spoori naelu, löögid või löögid, lill, lootele, seemned) teostavad taimede soo ja mitte-reprodutseerimisega seotud funktsioone ning tagada vormi olemasolu üldiselt, selle paljundamine ja ümberasustamine.
Orgaanite taimede lammutamine organitel tekkis nende struktuuri komplikatsioon järk-järgult taimemaailma arendamisprotsessis järk-järgult. Esimese maismaataimede - rhinofiidi või pylodiidi keha - ei levitatud juurtel ja lehtedel ning oli esindatud aksiaalsete kehade - kehaga - keha. Kuna taimed ulatuvad maale ja kohandavad neid elu- ja mulla keskkondade eluiga, muutus kehas muutus, mis viis elundite moodustumiseni.
Vetikad, seente ja samblike, keha ei erita organitele, vaid on esindatud Talomom või kihiline väga mitmekesine välimus.
Orgaanide moodustamisel tuvastatakse mõned tavalised mustrid. Kui kasvatate taimi, suureneb keha suurus ja kaal, rakkude jagunemine ja nende venitamine teatud suunas. Esimese etapi mis tahes neoplasma on orientatsioon rakuliste struktuuride kosmoses, s.o polaarsuse. Kõige kõrgemate seemnetaimede juures on polaarsus juba ZyGote'is leitud ja kujutatud väljatöötamine, kus moodustatakse kaks naeruväärset elundit: põgeneda ülemise neeruga ja juurega. Paljude ainete liikumine toimub polaarse juhtivate marsruutide puhul, st teatud suunas.
Teine muster on sümmeetria. See avaldub telje suhtes poolosade asukohas. On mitmeid sümmeetriatüüpe: radiaalne - saate veeta kaks (või rohkem) sümmeetriatasandit; Kahepoolne - ainult üks sümmeetriatasand; Samal ajal eristage kahekordset (seljaaju) ja ventraalset (kõhu) külge (näiteks lehed, samuti horisontaalselt kasvavad elundid, st plagiotroopse kasvu). Kasvav vertikaalselt - Orthotropic - omada radiaalset sümmeetriat.
Seoses põhikehade kohandamisega muutuvad nende funktsioonid uutele kindlatele tingimustele, mis põhjustavad nende modifikatsioone või metamorfoosi (mugulad, sibulad, selgrood, neerud, lilled jne). Taimede morfoloogias eristavad homoloogseid ja sarnaseid organeid. Homoloogilistel organitel on sama päritolu, kuid võib erineda vormis ja tehtud funktsioonides. Sarnased elundid teostavad samu funktsioone ja neil on sama välimus, kuid nende päritolu erinev.
Kõrgemate taimede organimede infektsiooni orienteeritud kasv (mis on reaktsioon välistegurite ühepoolsele mõjule (valgus, raskusastme, niiskus). Aksiaalsete kehade kasv valgusele määratletakse positiivsena (võrsed) ja negatiivse (peamise juure) fotootropismi. Orienteeritud Aksiaalsete taimede kasvu, mis on tingitud maise atraktsiooni jõu ühepoolsest tegevusest, on defineeritud kui geotropism. Positiivne geotroopia juure põhjustab oma suuna kasvu keskele, negatiivse geotropismi varre - keskelt.
Innekude põgenemine ja juur on saadaval embrüost, mis asub küpsemasse seemnes. Germinaalne põgenemine koosneb teljest (idase vars) ja seemnete lehed või cotyledons. Seemnete embrüo seemnete arv ulatub 1 kuni 10-12.
Embrüo telje lõpus on põgenemispunkt. See moodustab Meristeemi poolt ja on sageli kumerpind. See on kasvava või tipu koonus. Ülaosas põgenemise (APEX), juure lehed paigutatakse tuberkulli või rullide kujul pärast seemikud. Tavaliselt kasvavad lehed lehed kiiremini kui varre, samas kui noored voldikud katavad üksteist ja kasvupunkti, moodustades embrüo taglase.
Osa teljest, kus seemnete alused nimetatakse seemnetena; Ülejäänud gerstinaalse teljega, mis on gütüüledooni all, nimetatakse hüpokotüülile või stepper põlvele. Selle alumine ots läheb embrüonaalseks juurele, mis esitatakse seni ainult kasvava koonusega.
Seemne idanemise ajal hakkavad kõik tuumade elundid järk-järgult kasvama. Seemnest väljub idane juur. See tugevdab mulla noori taimset ja hakkab absorbeerima vett ja mineraalseid aineid selles lahustunud, andes peamise juure alguse. Põhijuure vahelise piiri ja varre piiripunkti nimetatakse juurekooki. Enamikus taimedes hakkab peamine juur harule ja teise, kolmanda ja kõrgemate tellimuste külgmised juured ilmuvad juurestiku moodustamiseni. Hypocotyle'il juurte vanade osade kohta varrel ja mõnikord lehtedel võib moodustada juurte vajutades.
Peaaegu samaaegselt assinaalsetest lastest (APEX) areneb esimese järjekorra tulistamine või peamine põgenemine, mis ka filiaalid, moodustades uue jookseb teise, kolmanda ja kõrgema tellimuse, mis toob kaasa peamise põgenemissüsteemi moodustamise.
Sarnaste (spaunas, kummutid, sõnajalad), nende keha (sprofüüdi) puhul arenevad nende keha (sporophyte) avastustest Zygootist. Sporofy elu algstaadiumis hoitakse väljapresside (gametofüütide) kudedes. Zygota arendab idu, mis koosneb väsivast põgenemisest ja root-poolusest
Niisiis, mis tahes kõrgema taime keha koosneb pildistamisest ja (va mossoidide) juurisüsteemide ehitatud korduvatest struktuuridest - võrsed ja juured.
Kõigis kõrgeima taime elundites on kolm koe süsteeme kate, juhtiv ja põhiline - pidevalt jätkuvalt organist pärit organist, mis peegeldab taimeorganismi terviklikkust. Esimene süsteem moodustab taimede välimise kaitsekate; Teine, mis hõlmab flora ja ksyleemi, sukeldatakse põhiliste kudede süsteemi. Peamine erinevus struktuuri juur, varre ja lehe määratakse erinevate jaotus nende süsteemide.
Esmase kasvu ajal, mis algab juurte ja varrede ülaosade lähedal, moodustub taime primaarse keha põhikomponendid. Primaarse ksüleem ja primaarne flolam ja nendega seotud parenhüümi kangad moodustavad keskse silindri või steli, varre ja taime primaarse keha juur. Seal on mitu tüüpi Stel.